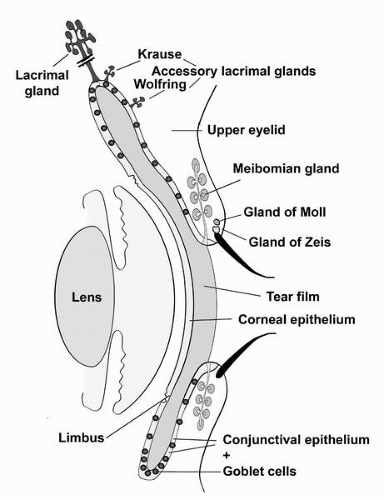
 FIGURE 1-1. Diagram demonstrating the ocular surface and adnexal tissues described in this chapter. The ocular surface tissues include the cornea, limbus, and conjunctiva; and the adnexa include the lids with eyelash cilia, the meibomian gland, the main lacrimal gland, the accessory lacrimal glands of Krause and Wolfring and the small glands, that empty into the cilia sheath—the glands of Zeiss and Moll.(see color image) |
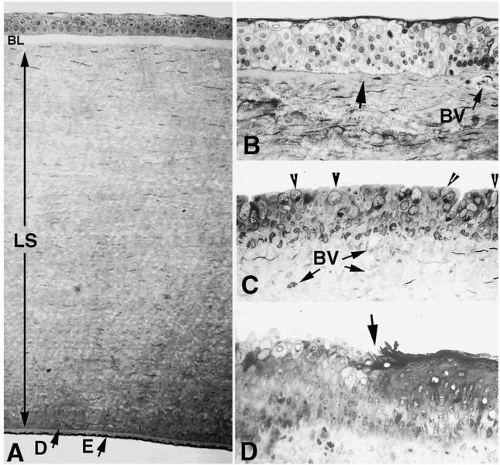 FIGURE 1-2. A: Histologic appearance of a full-thickness cross-section of human cornea shows the five- to seven-cell-layered epithelium, the three layers of the stroma, including Bowman’s layer (BL), the lamellar stroma with its resident keratocytes, which occupies over 90% of the corneal tissue (LS), and Descemet’s membrane (D), which is the thickened basement membrane of the corneal endothelium. The endothelium (E) is a low, cuboidal monolayer of cells that borders the anterior chamber. B: Light micrograph of anterior limbus showing the end of Bowman’s layer (arrow) at the juncture between cornea to the left and the first limbal blood vessel (BV) directly beneath the area of corneal epithelial stem cells. C: Micrograph of bulbar conjunctiva. Note numerous goblet cells (arrowheads) intercalated within the stratified epithelium as well as numerous blood vessels (BV) in the connective tissue. D: The junction (arrow) between nonkeratinized (left) and keratinized (right) epithelium of the epidermal tarsal and palpebral conjunctiva. Note numerous cells with the substantia propria below the epithelium. (Original magnifications: A, ×180; B, C, D, ×250.) |
potential danger. In addition to its unique functions over the translucent cornea, the epithelium carries out “routine” housekeeping functions common to any epithelium that borders the external environment. These include provision of a barrier to fluid loss and pathogen entrance, and resistance to abrasive pressure. A barrier to fluid loss is provided by tight junctions surrounding lateral membranes of apical cells, and a pathogen barrier is provided by membrane-spanning mucins at the apical surface. Resistance to abrasion requires that cells of the epithelium be specialized for tight adherence to one another and to their underlying extracellular matrix. Finally, its position adjacent to the outside world requires that the epithelium have a rapid and highly developed ability to respond to wounding.
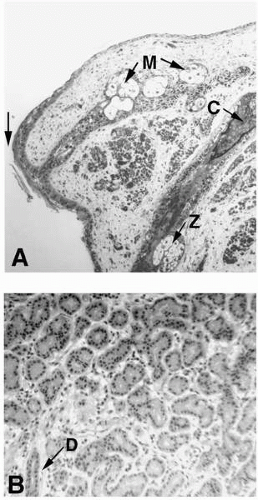 FIGURE 1-3. A: Low-power light micrographs of the lid margin showing the meibomian gland (M), a portion of the eyelash cilia (C), and its associated gland of Zeiss (Z). The juncture between conjunctival epithelium and epidermal epithelium is indicated by the arrow. B: Low-magnification micrograph of a section of human lacrimal gland. Acini are single cell layered. A duct (arrow) at D from the acini is shown running through connective tissue at lower left. (Original magnifications: A, ×120; B, ×220.) |
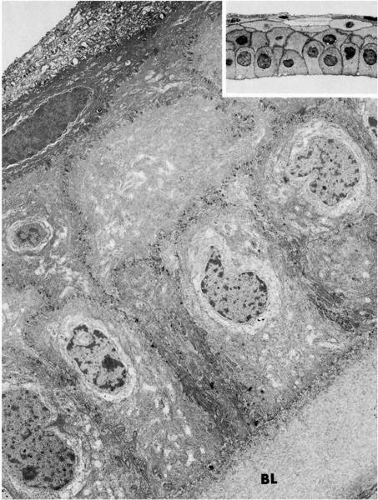 FIGURE 1-4. Low-magnification electron micrograph and light micrograph (inset) of the corneal epithelium and subjacent Bowman’s layer (BL). Note single layer of columnar basal cells, one to two layers of wing cells, and two to three layers of flattened squamous cells. (Original magnifications: main, ×4800; inset, ×750.) |
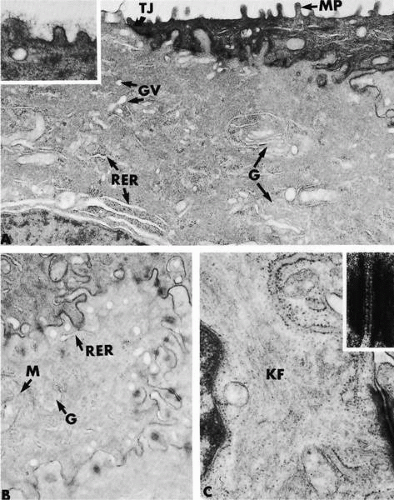 FIGURE 1-5. Electron micrographs demonstrating aspects of the ultrastructure of the corneal epithelium of apical cells (A plus inset) and wing cells (B and C plus inset). A: Apical and lateral borders of several adjacent apical cells are evident. Note microplicae (MP), region of tight junction (TJ), presence of Golgi apparatus (G), Golgi vesicles (GV), and rough endoplasmic reticulum (RER). The inset demonstrates a filamentous glycocalyx on the surface of the microplicae. B: Electron micrograph demonstrates the elaborate interdigitation of membranes of adjacent cells, characteristic of wing and squamous cells. A mitochondrion (M), Golgi apparatus (G), and rough endoplasmic reticulum (RER) are present. C: Higher-magnification electron micrograph demonstrates that the cytoplasm of epithelial cells is rich in keratin filaments (KF). A, B, and C all show the presence of the cell-cell anchoring junctions known as desmosomes, which are present along interdigitating cell membranes. A high-magnification section through a desmosome is shown in the inset of C. Desmosomes of corneal epithelia appear similar to those of all other stratified squamous epithelium. (Original magnifications: A, ×21,000, inset, ×51,000; B, ×21,000; C, ×42,000, inset, ×164,000.) |
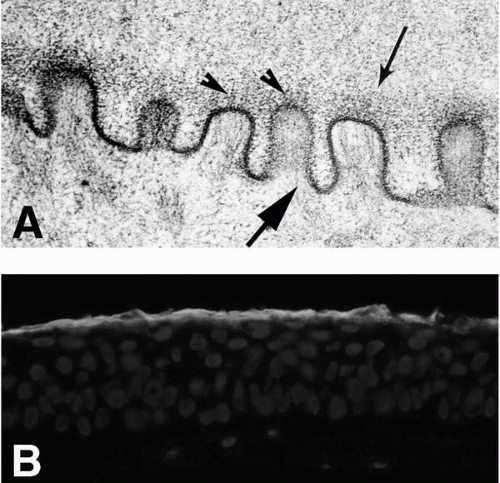 FIGURE 1-6. A: Electron micrograph of microplicae and glycocalyx (small arrow) on the surface of guinea pig conjunctiva. The membrane-associated mucins can be seen emanating from the tips of the microplicae (arrowheads) in the electron-dense glycocalyx. At least three different membrane-associated mucins are present in the human glycocalyx; these include MUC1, MUC4, and MUC16. The cytoplasmic domains of the mucins are believed to associate with actin filaments (large arrow), which extend toward the membrane where membrane-associated mucins insert. (From Nichols BA, Chiappino ML, Dawson CR. Demonstration of the mucous layer of the tear film by electron microscopy. Invest Ophthalmol Vis Sci 1985;26:464-473.) B: The immunohistochemical localization of one of the membrane-associated mucins, MUC16, is shown on a section of corneal epithelium. (Original magnifications: A, ×89,000; B, ×300.) |
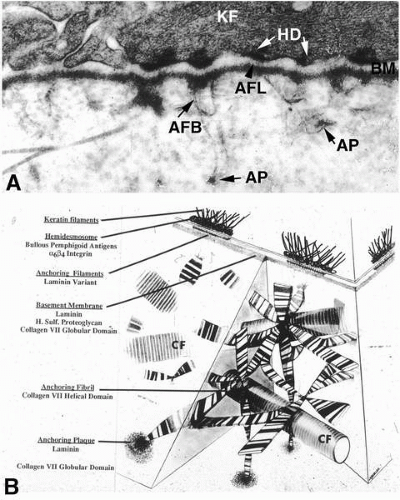 FIGURE 1-7. A: Electron micrograph of section through the epithelial-stromal junction region. The structures that are linked to form the epithelial anchoring complex can be seen. They include the hemidesmosome with keratin filaments (KF) inserting into the hemidesmosome plaque (HD). Extracellularly, anchoring filaments (AFL) can be seen in the lamina lucida zone of the basement membrane (BM). Anchoring fibrils (AFB) extend into the stroma at sites opposite the basement membrane from hemidesmosomes; anchoring fibrils insert into anchoring plaques (AP) distal from their insertion into the basement membrane. These plaques have the appearance of small bits of basement membrane. (Original magnification, ×65,000.) B: Diagram illustrating a three-dimensional view of the anchoring complex of the corneal epithelium. The column on the left lists the individual structures (underlined) of the linked complex with their known components underneath. The anchoring fibrils insert into the basement membrane opposite from hemidesmosomes. The cross-banded fibrils splay out among the collagen fibrils (CF), forming a three-dimensional network holding the epithelium tightly to the stroma. The anchoring fibrils terminate distally from the basement membrane in anchoring plaques. |
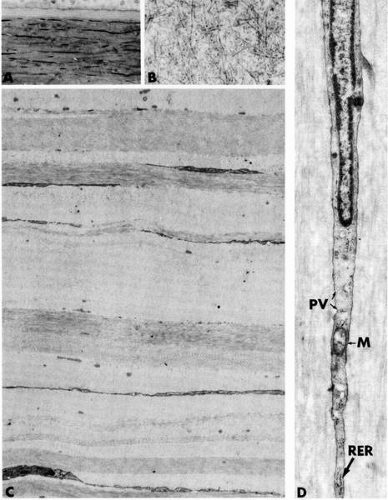 FIGURE 1-8. Micrographs depicting structural aspects of the corneal stroma. A: Light micrograph showing basal epithelial cells, subjacent Bowman’s layer, and the anterior lamellar stroma with its flattened and attenuated stromal fibroblasts, termed keratocytes. B: Electron micrograph of a region in Bowman’s layer demonstrating the random, felt-like interweaving of the collagen fibrils. C: Lower-power electron micrograph of the layered lamellar stroma. Note the presence of the keratocytes running between lamellae. D: Micrograph showing a segment of a flattened keratocyte. Rough endoplasmic reticulum (RER), a mitochondrion (M), and numerous pinocytic vesicles (PV) are present in the cytoplasm. (Original magnifications: A, ×300; B, ×31,000; C, ×4800; D, ×21,000.) |
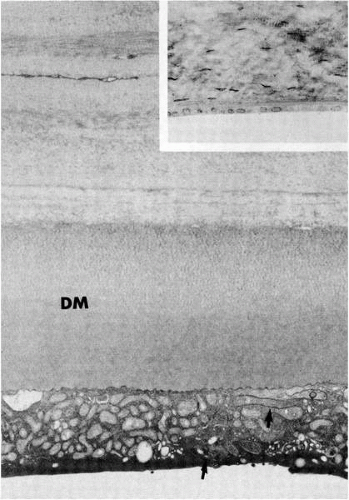 FIGURE 1-9. Micrographs of posterior corneal stroma, Descemet’s membrane (DM), and the corneal endothelium. The inset is a light micrograph section from a newborn with its comparatively thin Descemet’s membrane. The electron micrograph is of a section from an 18-year-old human. Note the two layers in Descemet’s membrane. The inner-banded layer was deposited by the endothelium during fetal life. The portion of the endothelial cell visible in the micrograph shows the presence of numerous mitochondria and an interdigitating lateral membrane (arrows). (Original magnifications: main, ×10,000; inset, ×300.) |
TABLE 1-1. CORNEAL EXTRACELLULAR MATRIX COMPONENTS | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||
glycoproteins. However, any glycoprotein with a collagenous domain is generally referred to as collagen. The remaining noncollagenous proteins in the stroma are subdivided into two major groups, glycoproteins and proteoglycans. Proteoglycans are a special class of glycoproteins that have glycosaminoglycan (GAG) side chains.
TABLE 1-2. COLLAGENS PRESENT IN CORNEA | ||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
|
proteins of lumican and decorin bind to collagen and limit the size of the fibril. The importance of proteoglycans in corneal stromal fibril formation is demonstrated in mice lacking lumican, which have corneal opacities as a result of irregular fibril size and spacing (64), and also in humans who have mutations in keratocan. This mutation results in corneal flattening (65).
synthesized by the epithelium in response to a wound [reviewed in Zieske (71)] (Table 1-1). These include laminin-5, entactin, collagen IV, and perlecan, which are normally present in the basement membrane. In addition, components not normally present are synthesized and deposited, including lumican, fibrin, and an unprocessed form of laminin-5. These components appear to be present in all types of wounds and make up a wound-healing surface that has, in the past, been termed a pseudomembrane, but is more properly termed a provisional matrix. Epithelial cells actually migrate on this provisional matrix after wounding, and several lines of evidence in animal models suggest that the matrix influences wound healing [reviewed in Zieske (71)]. For example, corneal epithelial cells in culture migrate at a faster rate on unprocessed laminin-5 than on the processed form seen in unwounded corneas. Stromal wound healing also appears to involve a provisional matrix. A number of extracellular matrix components not normally seen in the stroma have been localized after wounding, including fibronectin, fibrin, tenascin, collagen types IV and VII, and laminin-1 (Table 1-1). In addition, keratan sulfate proteoglycan levels decrease whereas chondroitin sulfate levels increase in the wounded stroma. In an experimental model, fibronectin and chondroitin sulfate stimulate fibroblast migration into a matrix. Thus, the provisional matrix may promote migration of fibroblasts into the wound area.
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


