Pathophysiology of the Vitreoretinal Interface
Yunyoung Kim
David J. Wilson
ANATOMY AND PHYSIOLOGY OF THE VITREORETINAL INTERFACE
It is not surprising that the junction of the vitreous and the retina is a critical interface for both physiologic and pathologic processes. At this site, there are numerous, abrupt transitions at the molecular, biochemical, cellular, and structural level. An appreciation of the features of molecular structure, histology, and gross anatomy of this interface is critical for understanding a diverse group of states including retinal development, retinopathy of prematurity, ocular trauma, retinovascular disease, retinal detachment, macular hole development, vitreous detachment, epiretinal membrane, and others.
The vitreoretinal interface is composed of the cortical vitreous, the internal limiting membrane (ILM; lamina) of the retina, and the innermost portion of the Mueller cells. It is easiest to consider this zone of tissue by initially appreciating the structure and function of the ILM, which represents the basement membrane of the Mueller cells. One can subsequently assess how the ILM is related to the cortical vitreous and the Mueller cell in a functional and structural sense.
Basement membranes are ubiquitous extracellular protein matrices that form a border between cells and their adjacent mesenchymal tissue. These membranes have multiple functions including structural cell support and attachment, interaction with cellular receptors, storage or repository site for growth factors and cytokines, tissue compartmentalization, and functioning as a molecular sieve to exclude proteins above a certain size. The interaction with cellular receptors has effects on cell shape, gene expression, cell migration and proliferation, and programmed cell death (1).
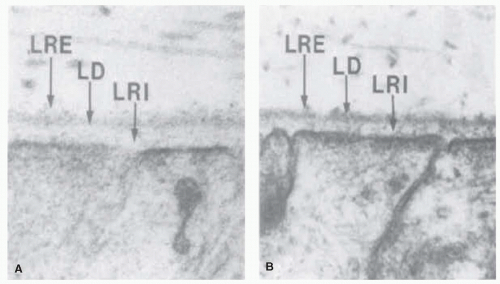 Figure 1-1. Electron micrographs of the medullary ray region of the rabbit eye showing the internal limiting membrane (ILM). A: Tissue fixed with glutaraldehyde and paraformaldehyde. The ILM is adjacent to the Mueller cell end feet and has three layers. One of these layers is electron dense (lamina densa [LD]) and is readily seen in the transmission electron micrograph (TEM). The other layers are electron transparent and are not evident in aldehyde-fixed material. The lamina rara interna (LRI) is between two electron-dense areas: the cytoplasm of the Mueller cell and the LD. The lamina rara externa (LRE) is on the vitreal face of the lamina densa and is an ill-defined layer, 5-10-nm thick. Collagen fibrils are 16-nm thick filaments without cross bands. (×96,000) B: Tissue fixed with Alcian blue. The LD is more electron dense after Alcian blue fixation, and material can be seen within the LRI (×62,000). (From Matsumoto B, Blanks JC, Ryan SJ. Topographic variations in the rabbit and primate internal limiting membrane. Invest Ophthalmol Vis Sci 1984;25:71-82, with permission.) |
Given the above list, it is simplistic and inaccurate to think of basement membranes as purely structural elements, but an appreciation of their structure is helpful in understanding the more dynamic aspects of this interface. The ultrastructure of the ILM contains three distinct areas: the lamina rara, the lamina densa, and the lamina rara interna (2) (Fig. 1-1). There are topographic variations in the ILM, and Foos (3) has divided these variations into three zones: basal, equatorial, and posterior. In addition to these three broad zones, there are specific features of the ILM of the fovea and the optic nerve.
In the basal zone, the ILM is uniformly thin, averaging 510 Angstroms (Å) in thickness. In this zone, the vitreous collagen fibrils have an orientation perpendicular to the surface of the ILM. The equatorial zone is characterized by a progressive and gradual thickening of the ILM, achieving an average thickness of 2600 Å. In this zone, the vitreous fibrils are oriented more tangentially to the surface of the ILM. In the posterior zone, the ILM has an average thickness of 25,000 Å, and the retinal surface is markedly uneven, corresponding to the undulating surface of the Mueller cells. In the posterior zone, the overlying vitreous fibrils continue to have an orientation tangential to the surface of the ILM.
One distinction between these three zones is the relative abundance of Mueller cell attachment plaques. These plaques represent condensations of the fibrils of the Mueller cell cytoskeleton. Attachment plaques were noted to be more
abundant in the basal zone and virtually absent in the posterior zone. Foos (3) attributed the topographical differences of vitreous fibril orientation and attachment plaque density to the variation in the amount of vitreous traction in the different zones.
abundant in the basal zone and virtually absent in the posterior zone. Foos (3) attributed the topographical differences of vitreous fibril orientation and attachment plaque density to the variation in the amount of vitreous traction in the different zones.
The ILM of the fovea is distinct from the remainder of the posterior zone, in that it is much thinner, starting at the clivus, diminishing to a thickness of 200 Å at the foveola. Mueller cell attachment plaques also reappear in this same location. At the optic nerve head margin, the thick ILM of the posterior zone abruptly diminishes to a thickness of 450 Å.
The ILM, like other basement membranes, is composed of several collagens, laminins, proteoglycans, calcium-binding proteins, and other structural and adhesive proteins (1) (Table 1-1). Type IV collagen provides a highly cross-linked structural framework that maintains mechanical stability. A second network of laminins is more variable in composition between tissue and also may vary within the same tissue (1). There have been only limited studies of the specific laminin and other protein/proteoglycan content of the ILM (4, 5, 6 and 7). Studies in other tissues have implicated laminin binding sites as providing the adhesion of the basement membrane to the underlying cells (1,8), possibly through alpha-dystroglycan linkage of the basement membrane to the actin skeleton of the underlying cell. Although ultrastructural studies initially suggested the vitreous collagen inserted into the collagenous network of the ILM, more recent studies suggest that this adhesion is the result of noncovalent binding of proteoglycan moieties to the vitreous and basement membrane collagens (7). Hence, the adhesion of the vitreous to the retina is not maintained by a collagenous network but rather by noncovalent binding of laminins and proteoglycan constituents of the vitreoretinal interface. This mechanism of adhesion can be affected by various degenerative and pathologic states as we will see later in this chapter.
In addition to its mechanical and structural features, the ILM is capable of playing a dynamic role by regulation of cell binding and of the local cytokine microenvironment. Basement membranes contain multiple proteins, that when bound by cell surface receptors initiate intracellular signaling pathways that alter cellular behavior. During basement membrane remodeling, exposure of these proteins can lead to cellular activities that promote tissue repair such as recruitment of immune cells and activation of fibroblasts. ILM also is a repository for vascular endothelial growth factor (7). Following injury, or as a result of disease, vascular endothelial growth factor and other stored cytokines may impact the microenvironment for angiogenesis. Anti-angiogenic fragments of basement membrane collagen released during remodeling of the ILM (endostatin, tumstatin, and arresten) may also play a role in the angiogenic response at the vitreoretinal interface (9).
TABLE 1-1 REPRESENTATIVE BASEMENT MEMBRANE PROTEINS | ||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ||||||||||||||||||||||||||||||||||||||||||||||||
Changes that occur in the retina and the vitreous often have their clinical manifestations at the vitreoretinal interface. These changes may be on the basis of mechanical injury, metabolic processes, or age-related degeneration. Because the vitreoretinal interface is the junction between the relatively fluid vitreous and the retina, the manifestations of a wide variety of diseases occur at this location.
Age-Related Changes in the Vitreoretinal Interface
Age-related and other degenerative changes in the vitreous commonly lead to separation of the vitreous from the ILM. The molecular basis for this degeneration has not been fully elucidated, but recent studies have found that there is fragmentation of vitreous collagen that leads to the formation of pockets of fluid within the normally homogeneous matrix of collagen separated by a proteoglycan matrix (10,11). This degeneration may be catalyzed by endogenous enzymes including plasmin and other matrix metalloproteinases (12, 13 and 14).
The availability of optical coherence tomography (OCT) has provided great insight into the process of separation of the vitreous from the ILM, in particular how this separation is related to the pathophysiology of many common retinal conditions (15). Uchino and coworkers (16




Stay updated, free articles. Join our Telegram channel
Full access? Get Clinical Tree
