Fig. 29.1
Target azimuth tuning functions of one IC neuron under altered-cue and standard conditions for a single source (a) and with a central interferer (b). (c) Target ITD tuning functions from a cross-correlation model using a cochlear filter with center frequency, f center = 3.19 kHz. Interferer location (solid triangle); ITD equal to one period of f center (open triangles)
The restriction of rate suppression and enhancement to high-BF neurons suggests the involvement of ILDs or envelope ITDs, as these are the binaural cues to which high-BF neurons are sensitive (Delgutte et al. 1995; Joris 2003). To identify the relevant cues, we compared azimuth tuning functions between altered-cue and standard conditions. For the high-BF neuron in Fig. 29.1a, the single-source tuning function in the fixed-ITD condition resembles that in the standard condition, while the tuning function in the ITD-only condition is nearly flat, suggesting that tuning is primarily determined by ILD. In contrast, in the presence of a central interferer (Fig. 29.1b), the fixed-ITD tuning function no longer resembles the standard tuning function, while the ITD-only tuning function shows rate enhancement similar to that observed in the standard condition. Therefore, the rate enhancement with target separation from a central interferer was dependent on sensitivity to ITD, although the single-source tuning function was primarily dependent on ILD. This neuron was representative in that rate suppression or enhancement was consistently observed in the ITD-only condition but not the fixed-ITD condition.
Rate suppression or enhancement can be explained by a cross-correlation model operating on cochlea-induced envelopes of the ear input signals. In this model, left and right sound stimuli were band-pass filtered (0.25-oct bandwidth) to mimic cochlear processing, and the normalized correlation (Trahiotis et al. 2005) was computed between the Hilbert envelopes of the left and right band-pass-filtered waveforms. As the ITD of a broadband source is varied, the interaural cross-correlation coefficient (IACC) decreases slowly (Fig. 29.1c, dashed line) because the envelopes being cross-correlated only contain low frequencies. However, when a central interferer is introduced (Fig. 29.1c, solid line), the IACC changes more rapidly with target ITD, oscillating at the period of the filter center frequency (3.19 kHz in Fig. 29.1c). While the envelopes in this case are still limited to low frequencies, the rapid oscillation occurs because the waveform interactions between the target and interferer that determine the mixture envelope are dependent on the temporal fine structure after cochlear filtering. The behavior of IC neurons that are sensitive to envelope ITD is consistent with model predictions. While the firing rates of IC neurons typically change little when a single source moves by 15° (Fig. 29.1a, dashed line), the azimuth tuning functions with a central interferer show sharp changes in firing rate as the cochlea-induced envelopes become strongly decorrelated when the target is separated by 15° from the interferer.
Best et al. (2004) showed that the perceptual segregation performance for two broadband noise sources (with one fixed at 0°) remains good when the stimuli are high-pass filtered but is degraded in the fixed-ITD condition, suggesting the use of envelope ITD for segregation. The sharp rate suppression or enhancement seen in high-BF IC neurons is a possible neural correlate of this ability. To test this idea, we calculated a neural detection threshold for the spatial separation of the target from the central interferer for each high-BF neuron in our sample (Fig. 29.2a, b). The median detection threshold across the population of high-BF neurons was small (<8°) and nearly the same for ITD-only and standard conditions but was 4 times larger for the fixed-ITD condition (Fig. 29.2c). Thus, the sensitivity of IC neurons to envelope decorrelation plays a key role in the neural detection of source separation for high-pass sounds.
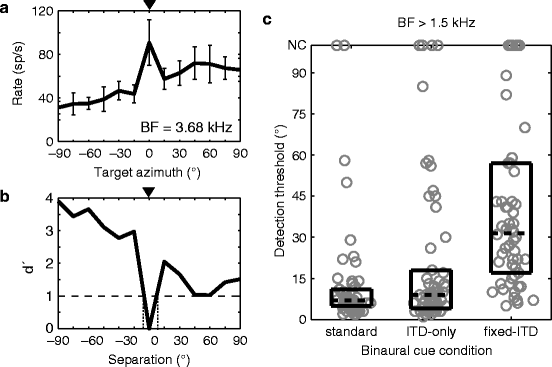
Fig. 29.2




Neural signal detection. (a) Target azimuth tuning function in the presence of a central interferer for a single IC neuron (bars: 1 SD). Triangle indicates interferer location. (b) d′ calculated from the data in (a) showing the detection thresholds (dotted lines) where d′ = 1 (dashed line). (c) Distribution of detection thresholds across high-BF neurons (N = 58) showing medians (dashed lines) and interquartile ranges (boxes)
Stay updated, free articles. Join our Telegram channel
Full access? Get Clinical Tree
