Morphology and Pathologic Response in Corneal and Conjunctival Disease
Kenneth R. Kenyon
Emiliano Ghinelli
Henrique V. Chaves
The cornea is composed of relatively few components and includes three different cell types (epithelium, keratocytes, and endothelium) plus an extracellular matrix of collagen and glycosaminoglycans. Thus it is appropriate to consider corneal morphology and pathology in terms of the three major anatomic and functional units formed by these cells and matrix: the epithelium and its basement membrane, the keratocytes and the stromal collagen, and the endothelium and Descemet’s membrane (1,2).
Because normal corneal morphology is discussed elsewhere (Chapter 1), this chapter surveys the basic principles of normal wound healing, the response to specific insults (such as chemical injury, bacterial infection, and surgical procedures), and the response during selected diseases.
EPITHELIUM AND BASEMENT MEMBRANE
Normal Wound Healing
Subsequent to a full-thickness wound of the corneal epithelium, the damaged cells adjacent to the wound edge lose their surface microvilli (3, 4, 5). Within 1 hour, the basal cells begin to flatten as intermediate and superficial cells reduce their desmosomal junctions and glycogen storage, evidencing their impending movement.
Fibronectin, the glycoprotein widely involved in cell-to-cell and cell-to-substrate interactions, is deposited on the denuded corneal surface, along with fibrinogen and fibrin (6). Neutrophils arrive through the tear film, initiating the process of debriding cellular remnants. At 6 hours postwounding, the epithelial cells initiate wound closure by sliding into the area of defect at a rate of approximately 0.75 mm/min (7,8). At the same time, basal cells distant from the wound begin to undergo mitosis.
The epithelial sliding process is quite active for 15 hours, as the leading cells extend ruffled pseudopods whose motility depends on the actin filaments of the cytoskeleton (9). The cytoskeletal filaments may also be involved in cell adhesion because they insert into the electron-dense inner aspect of the hemidesmosome attachment plaques.
The moment the defect is closed, a contact inhibition message evokes the cessation of cell movement and the alteration of cellular configuration, causing these migrating squamous cells to regain the cuboidal configuration of basal cells. At the same time, DNA synthesis begins. Between 24 and 48 hours after wounding, epithelial proliferation in the wound, now at its peak, forms an epithelial plug, and all neutrophils vanish. By 3 to 4 days after wounding, the epithelial plug regresses and mitotic figures first appear in the wound (10) (Fig. 4-1).
If the epithelial basement membrane remains healthy, the recovering epithelial cells are able to use it for tight adhesion to the underlying stroma, as is evidenced both morphologically by the reappearance of hemidesmosomes on day 2 after injury and clinically by tight adhesion shortly thereafter (11).
In the event of basement membrane damage, however, new basement membrane complexes, comprising short, discontinuous segments of basement membrane material with associated hemidesmosomes and anchoring fibrils, begin to reform within 5 to 7 days after injury (12). Depending on the severity of basement membrane damage, as long as 6 to 8 weeks may be required for complete basement membrane reconstruction. This process is closely paralleled by delayed epithelium-to-stroma adhesion until the intact basement membrane has been restored (13). After regression of the epithelial plug, mitoses appear in the wound base, and soon the overlying new epithelium becomes even with the adjacent old epithelium. The regenerating epithelium is important in activating the underlying keratocytes because they fail to transform into fibroblasts in the absence of overlying epithelium (14,15). The damaged epithelium may also be involved in the recruitment of neutrophils into the wound area. Moreover, neutrophils and mononuclear cells can delay the closure of the epithelial wound, such that continuing inflammation may foster the persistence of a corneal epithelial defect (16). A crucial role in corneal epithelial wound healing is also played by hormones, biochemical messengers, and pharmacologic agents. The classic studies of Friedenwald and Bushke (17) indicated that influences on corneal epithelial healing were adrenergically mediated because topically administered epinephrine reduced epithelial cell locomotion, mitosis, and wound healing. In addition, second messengers, including adenosine 3′,5′-cyclic adenosine monophosphate (cAMP) and guanosine 3′,5′-cyclic guanosine monophosphate (cGMP), have been implicated in corneal epithelial cell metabolic control (18). The proliferation of corneal epithelial cells is regulated by a bidirectional control process characterized by an adrenergic cAMP-dependent “off” response, and a cholinergic, muscarinic cGMP-dependent “on” response. Exogenous substances that raise intracellular cAMP levels such as isoproterenol or prostaglandin-E1 shut off epithelial mitosis, and carbamylcholine and acetylcholine (ACh) raise intranuclear cGMP levels and increase mitosis by specific, regulatory stimulation of RNA polymerase II activity (19). This mechanism explains the transitory mitotic suppression induced by superficial corneal wounding (interruption of adrenergic fibers), as well as the permanent depression of epithelial mitosis associated with decreased intracellular ACh levels after total corneal denervation, resulting in neurotrophic keratitis (20, 21, 22, 23).
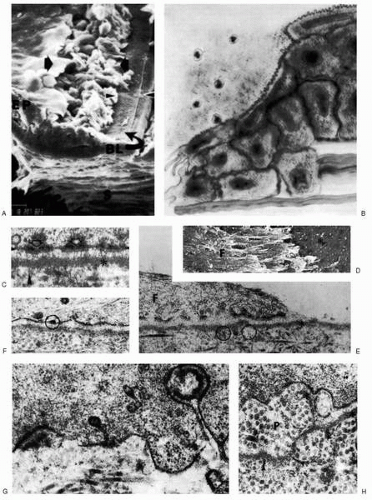 FIGURE 4-1. Normal epithelial wound healing. A: Scanning electron micrograph immediately after trephine incision of epithelium and stroma, with the wound margin seen at the right. Immediately adjacent to the wound margin, an exposed zone of Bowman’s layer (BL) appears intact (between the arrowheads) but is devoid of epithelial cells. Farther from the wound margin, an area of disrupted epithelial cells is evident (between the arrows), and more distant from the wound edge, the epithelium (EP) appears unaffected. S, stroma (×600). B: Diagram of the epithelium 15 to 72 hours after wounding. Epithelial sliding is active as filopodia extend from the crest and edges of ruffled epithelial cells into the defect area. Intracellular spaces are distended. As the base of the wound is covered, the cells increase in height and begin again to resemble basal cells. Cellular organelles increase in size and complexity, and villous specializations of the free surface membrane begin to reappear. (A, B from Binder PS, et al. Corneal anatomy and wound healing. In: Transactions of the New Orleans Academy of Ophthalmology. St. Louis: Mosby, 1980:1-35, with permission.) C: Transmission electron micrograph at high magnification shows basement membrane attachment complexes to comprise cytoplasmic filaments within basal epithelial cells, converging at hemidesmosomes (arrows), penetrating the plasma membrane (small arrowheads), bridging the lamina lucida between the plasma membrane and basement membrane (asterisk), and establishing continuity with anchoring fibrils (large arrowhead) in Bowman’s layer (×100,000). D: Scanning electron micrograph of rabbit cornea immediately after surgery reveals filamentous cellular debris (F) adherent to exposed and intact basement membrane (asterisk; ×1500). E: Transmission electron micrograph of rabbit cornea corresponding to D. The basal cells have been ruptured transversely, leaving masses of filamentous debris (F) overlying an intact basement membrane (asterisk) with well-developed anchoring fibrils (circled; ×30,000). F: Transmission electron micrograph of rabbit cornea 3 days after mechanical scraping of the epithelium. Newly migrating epithelium has come to rest on the original basement membrane, and hemidesmosomal attachment sites (circled) have formed (×40,000). G: Transmission electron micrograph of rabbit cornea 7 days after superficial keratectomy illustrates the beginnings of the attachment complex reconstruction as short, discontinuous segments of basement membrane (asterisk) with attendant hemidesmosomes along infoldings of the basal cell membrane (×40,000). H: Transmission electron micrograph of rabbit cornea 30 days after superficial keratectomy shows areas of irregularity and duplication of basement membrane (asterisks), separated from basal cell membrane by fibrillar collagenous pannus (P), as evidence of probable epithelial erosion or edema. Numerous anchoring fibrils (arrowheads) are evident (×40,000). |
Another important modulator of mitotic rate in corneal epithelium is human epidermal growth factor (hEGF). Classic works performed by Frati and coworkers (24) and by Savage and Cohen (25) independently showed that hEGF applied to wounded rabbit corneas stimulates reepithelialization by a transient hyperplasia of epithelial cell layers. Soong and coworkers (26) found that acceleration of corneal wound healing by hEGF is entirely due to stimulation of cellular mitosis, not to increased cell motility. Also, Kitazawa and coworkers showed that topical application of hEGF in rabbits induces a dose-dependent high rate of epithelial replication, particularly near the limbal region, with a significant increase in the DNA content in regenerating epithelium over 24 to 48 hours.
Transforming growth factor-β (TGF-β) is a multifunctional peptide that modulates cell proliferation and differentiation in many cell types and accelerates tissue repair response (27,28). In vivo studies have shown that TGF-β present in fibroblasts, macrophages, and possibly the cell membrane may accelerate stromal wound healing, induce neovascularization, and decrease plasminogen activator (PA) activity, resulting in the prevention of degradation of extracellular matrix. However, in the absence of retinoic acid, the effect of TGF-β is more prominent, suggesting
that TGF-β plays a more important role in the healing of vitamin A-deficient corneas than in normal corneas (29) (Fig. 4-2).
that TGF-β plays a more important role in the healing of vitamin A-deficient corneas than in normal corneas (29) (Fig. 4-2).
 FIGURE 4-2. Immunohistochemical staining of transforming growth factor-beta (TGF-β) in the vitamin A-deficient rat cornea. At 24 hours after superficial mechanical injury (A) a positive reaction for TGF-β is evident as well as the formation of a pseudomembrane (asterisks) in the central epithelial defect and anterior stroma. At higher magnification (B) many infiltrating inflammatory cells and keratocytes show a positive reaction (A ×87; B ×220). |
Nishida and coworkers (30) also showed that hyaluronan (hyaluronic acid) stimulates corneal epithelial migration by mechanisms different from those of fibronectin and hEGF. Further studies are necessary to prove that beyond its utility as a viscoelastic substance in ophthalmic surgeries, hyaluronan with its biologic signal to cells could be therapeutically promoting corneal epithelial wound healing.
Several studies have shown that fibronectin, a high-molecular-weight glycoprotein found in plasma, on cell surfaces, and in extracellular matrices, is an important factor in adhesion, chemotaxis, tissue repair, and phagocytic activity of keratocytes (31, 32, 33, 34, 35, 36, 37, 38, 39). After abrasion of the corneal epithelium, fibronectin is detected in the anterior margin of the denuded stroma in areas where the epithelium has not healed 18 hours after the injury, but not 40 hours later (40). Recent experimental studies suggest that fibronectin might be important in the healing of corneal epithelial wounds (6,41, 42, 43, 44, 45, 46), although other investigators could not prove the clinical usefulness of fibronectin to promote wound healing of corneal epithelium (47,48).
The proteinases plasmin and PA have been detected in ocular tissues (49,50), and Berman and Wang suggest their involvement in ocular surface disorders involving sterile corneal ulceration (51,52).
Plasmin is able to degrade fibronectin and laminin (53, 54, 55, 56), both of which are important in cell sliding during corneal wound healing.
Recent studies have shown that urokinase-like PA (uPA) but not tissue-type PA is present at the leading edge of migrating corneal epithelium both after abrasion and after alkali injuries (57, 58, 59). The abnormally prolonged presence of enzymatically active uPA at the leading epithelium edge after burn results in a persistent corneal epithelial defect. Therefore, the regulation of uPA activity at the leading edge of corneal epithelium after injury might be useful therapeutically in the healing of epithelial defects and the prevention of stromal ulceration.
Recently, the mechanisms of action of EGF and other factors, molecules, and receptors have been summarized and a mechanism proposed for the different and specific moments of the corneal wound healing process (60). During the cell migration phase of the corneal wound repair, many different molecules are considered key actors that variously affect the early phase of epithelial renewal. They include fibroblast growth factor (FGF), TGF, keratinocyte growth factor, hepatocyte growth factor, platelet-derived growth factor, insulin-like growth factor, interleukin (IL)-1 and IL-6, tumor necrosis factor-α, endothelin-1, fibronectin, retinoids, CD44, hyaluronan receptor, cytokeratins 3 and 12 (K3 and K12), amyloid precursor protein, and chondroitin sulfate proteoglycans (60). Some peptides like nerve growth factor and substance P also play a role in the process, providing evidence that the corneal epithelium is richly innervated and responsive to neuropeptides (60,61).
Response to Limited Ocular Surface Injury
The healing rate of the ocular surface epithelium varies with the extent as well as the intensity of the injury (Fig. 4-3). Limited mechanical abrasion and mild chemical or thermal burns of the central cornea usually heal rapidly by the mechanism detailed in the previous section. In the rabbit, regenerated corneal epithelium shows restoration of its biochemical components, including glycogen and glycolytic enzymes, as soon as healing is completed, and this
corresponds to a normal clinical and morphologic appearance as well (62,63). However, with more severe injury of the corneal surface, involving damage to the basement membrane complexes of the basal epithelial cells and the anterior stroma, recovery may be slower. For example, extreme chemical burns involving both the corneal and conjunctival epithelia heal slowly with recurrent epithelial erosions, scarring, and neovascularization; this pattern of healing is related in part to the intrinsically different regenerative capacities of the corneal and conjunctival epithelial cells, especially after widespread destruction of the limbal stem cells.
corresponds to a normal clinical and morphologic appearance as well (62,63). However, with more severe injury of the corneal surface, involving damage to the basement membrane complexes of the basal epithelial cells and the anterior stroma, recovery may be slower. For example, extreme chemical burns involving both the corneal and conjunctival epithelia heal slowly with recurrent epithelial erosions, scarring, and neovascularization; this pattern of healing is related in part to the intrinsically different regenerative capacities of the corneal and conjunctival epithelial cells, especially after widespread destruction of the limbal stem cells.
Compromise of the epithelial cells and their surface characteristics is clinically evident as surface irregularity and superficial punctate keratitis (corresponding to microscopic epithelial defects) and also as loss of cell surface microplicae. Although basement membrane damage can also contribute to the formation of persistent epithelial defects or recurrent erosions, as described later, these problems are fortunately unusual. Thus, for example, even after a relatively extensive acid burn of the human cornea and bulbar conjunctiva, epithelial recovery is rapid and secure (64). As shown in Fig. 4-3, the denatured corneal epithelium demonstrates nuclear pyknosis and cytoplasmic shrinkage. However, transmission electron microscopy fails to disclose any basement membrane material adherent to these devitalized basal epithelial cells, and the functional integrity of the original basement membrane is clinically attested by tight adhesion of the regenerated epithelium, which rapidly resurfaces the entire cornea and involved bulbar conjunctiva. Because of limited acid-induced denaturation of stromal collagen and ground substance, topical steroids can be liberally applied to inhibit inflammation.
Should, however, chemical penetration be more severe, then subepithelial fibroblastic proliferation may result in superficial stromal scarring in the form of a fibrocellular pannus with or without neovascularization. This pannus comprises randomly arrayed type II collagen fibrils of variable sizes and is the product of keratocytes that have been stimulated to transform into fibroblasts. Such a pannus may accumulate between the epithelium and Bowman’s layer or, more commonly, may develop in the anterior stroma with minimal detriment to corneal clarity and vision (65, 66, 67).
Response to Extensive Ocular Surface Injury
When an injury involves a large area of the ocular surface, a different pattern of healing occurs (Fig. 4-4). Subsequent to Thoft’s initial hypothesis of the conjunctiva’s role in the maintenance of the normal corneal epithelial cell mass, studies have shown that the stem cells (cells with great potential for clonogenic cell division and with responsibility for cell replacement and tissue regeneration) of the corneal epithelium are localized in the basal cell layer of the limbus (68, 69, 70, 71, 72) (Chapter 1). In fact, when the wound involves the entire cornea and extends beyond the limbus, thereby destroying the stem cells, the corneal surface must be recovered with epithelium of conjunctival origin, suggesting that the limbal epithelium also may be a barrier between corneal and conjunctival epithelia (73). Further experimental studies have also shown that total or partial surgical removal of limbal stem cells contributes to the triad of conjunctival epithelial ingrowth, corneal vascularization, and delayed healing with recurrent erosion (74,75). In this situation, the obligate transformation of conjunctival cells into corneal epithelium is prolonged functionally, morphologically, and biochemically (62, 63, 64). Such eyes face extended clinical morbidity because they are beset with chronic inflammation, persistent epithelial defects, recurrent erosions, stromal neovascularization, and sterile ulceration. Epithelial cells at the margin of persistent defects often display an elevated border, having undergone an arrest of both migratory and mitotic activity. Nor is there synthesis of the basement membrane material that may have been chemically denatured by the injury (76).
The overgrowth of conjunctival epithelium onto the cornea is also marked by the presence of goblet cells. Such retention of goblet cells on the cornea has been observed to correspond very closely to areas where the stroma has become vascularized, perhaps indicative of the persistently altered biochemical state and metabolic needs of conjunctivally derived epithelial cells (63,77). Somewhat paradoxically, as has been observed in chemically burned human eyes, not only are goblet cells numerous and morphologically normal (78), but non-goblet epithelial cells appear to be increasingly involved in mucus secretion (79). Hence, in contrast to previous interpretations, mucus deficiency need not be invoked as a significant contributor to the ocular surface abnormalities of the chemically burned eye (Fig. 4-5).
The conjunctiva-derived epithelium also has a dramatic effect on the response of the corneal stroma to subsequent injury; a rabbit cornea covered by epithelium of conjunctival origin demonstrates a marked stromal neovascular response to subsequent wounding (80). These findings explain why the response to major destruction of the limbal zone includes corneal vascularization and conjunctivalization, as well as why limbal transplantation is more effective than conjunctival transplantation in restoring a severely damaged corneal surface (81,82).
The whole anterior ocular surface is accessible to growth factors, especially neurotrophins, which are produced and released by the lacrimal gland into the tear film (83). These important growth factors can presumably be used by the conjunctival epithelium and possibly also the goblet cells because these cells show a plethora of neurotrophin receptors (84). Cell culture studies of human limbal epithelium have shown high rates of cell multiplication compared with the central cornea and conjunctiva, and have also demonstrated the ability of limbal cells to attach and form a stratified epithelium when suitably grafted (85) (Fig. 4-6). Recently, autologous fibrin-cultured limbal stem cells have been shown permanently to restore the corneal surface of patients with total limbal stem cell deficiency (86). These results again support the concept of limbal location of stem cells in the cornea, and suggest that monolayers of cultured epithelial cells may be useful as autografts for repair of a damaged ocular surface.
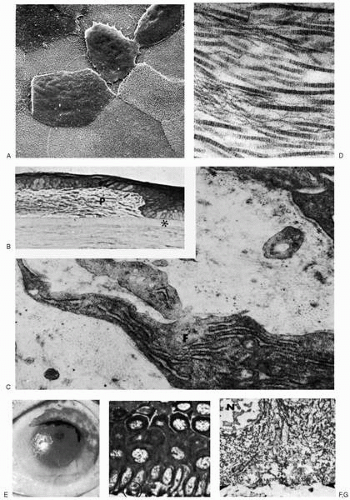 FIGURE 4-3. Healing response to mild ocular surface injury. A: Scanning electron micrograph of monkey cornea after thermal cauterization illustrates the morphology of superficial punctate keratitis; individual epithelial cells have lost surface membrane infoldings and are desquamating. Underlying and surrounding normal-appearing epithelium exhibits surface microplicae (×1000). B: Phase-contrast photomicrograph of human cornea 1 year after penetrating injury demonstrates proliferation of fibrocellular pannus (P) between the epithelium and an intact Bowman’s layer (asterisk; paraphenylenediamine, ×500). C: Transmission electron micrograph of a corresponding area reveals the basement membrane (asterisks) to be complete except for focal discontinuity. Multiple fibroblastic cells (F) in collagenous pannus appear synthetically active, as evidenced by abundant rough endoplasmic reticulum (×42,000). D: At higher magnification, a transmission electron micrograph reveals collagenous components of pannus to be randomly organized, banded fibrils of variable large diameter (up to 500 Å), with fine filaments interspersed (×62,000). E: A 33-year-old woman sustained an extensive burn by weak acid of nearly the total ocular surface epithelium. Anatomic and visual recovery was excellent after vigorous topical steroid and antibiotic therapy. F: Phase-contrast photomicrograph of debrided corneal epithelium (48 hours after injury) of the eye shown in E demonstrates preservation of the epithelial cell organization, despite cytoplasmic coagulation (paraphenylenediamine, ×1000). G: Transmission electron micrograph of the area circled in F shows nuclear (N) and cytoplasmic coagulation of basal epithelial cells. The basal plasma membrane (between the arrowheads) is notably devoid of basement membrane, which is presumed to have remained attached to Bowman’s layer (×5000). |
Keratinization: Xerophthalmia
Xerophthalmia is a highly clinically relevant example of ocular surface keratinization. Xerophthalmia is the term used to describe the irregular, lusterless, poorly wettable surface of the conjunctiva and cornea associated with vitamin A deficiency. Bitot’s spots, the triangular, foamy, keratotic areas of interpalpebral bulbar conjunctiva adjacent to the limbus, are also sometimes correlated with vitamin A deficiency. In humans, biopsy specimens of xerotic conjunctiva and cornea show remarkable acanthosis and hyperkeratosis with a prominent granular cell layer containing keratofibrils and keratohyalin granules (87,88) (Fig. 4-7A-D). Frequently, pleomorphic gram-positive bacilli, presumably diphtheroids, are clustered about the superficial keratin debris. Electron microscopy shows that the superficial cell layers appear cornified and have lost almost all recognizable organelles except extremely compacted keratofibrils (Fig. 4-8). The surface plasma membrane has greatly reduced microplicae, as confirmed by scanning electron microscopy. If vitamin A therapy is effective, clinical and histologic normalization begins within 1 to 2 weeks, although conjunctival goblet cells are initially evident only at 1 month, and may require 2 to 3 months or longer for complete restoration (87). The effect of topical drugs and preservatives on the tears and the corneal epithelium during xerophthalmia due to dry eye represents a very important issue in the pathogenesis of this condition, because the same topical agent used to treat a compromised ocular surface epithelium can also impede the healing of a wounded corneal epithelium (89).
Cicatrization: Pemphigoid
Ocular cicatricial pemphigoid is a slowly progressive, chronic condition primarily affecting the mucosa of the mouth, nose, pharynx, and conjunctiva with formation of bullae, shrinkage, and scarring (Chapter 20). Cutaneous lesions may involve the face and scalp. These bullous lesions occur between the basal cells and basement membrane, and in a fully developed blister the basement membrane is usually destroyed. In many patients with clinically active disease, immunofluorescence studies show fixed immunoglobulins in the basement membrane zone (BMZ) of involved skin or mucosa as well as circulating basement membrane antibodies (90,91). The production of autoantibody reactive with a component in the lamina lucida of the conjunctival epithelial BMZ activates the complement pathway, leading to monocyte and mast cell recruitment and degranulation (immunoglobulin E-independent). This in turn amplifies the reaction with vascular damage, fibroblast proliferation, and collagen synthesis, plus increased basal epithelial cell mitosis and squamous metaplasia. These immune deposits are bound at sites where the earliest pathologic lesions are identified, and the circulating autoantibodies reactive with the lamina lucida of the BMZ play a central role in lesion production (92). With progression of conjunctival involvement, connective tissue proliferation and symblepharon result. In extreme cases, xerosis, keratinization of the conjunctiva, and epidermalization of the cornea develop (Fig. 4-7E-I).
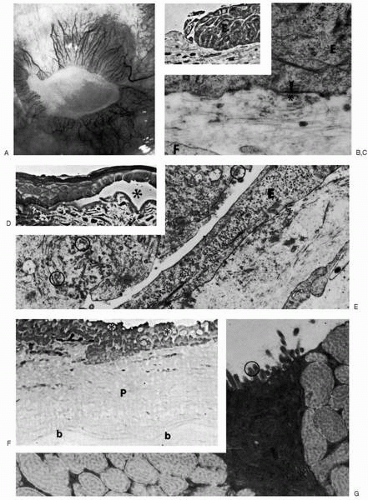 FIGURE 4-4. Healing response to severe ocular surface injury. A: A 43-year-old industrial worker is seen 1 year after acid and alkali injury to the entire ocular surface, with a central persistent epithelial defect surrounded by vascularized conjunctival overgrowth. B: Phase-contrast photomicrograph of monkey cornea 2 weeks after thermal cauterization shows multilayered, amitotic epithelium (E) at the edge of a persistent epithelial defect. Severe inflammatory and fibrocytic cells appear in the superficial stroma (paraphenylenediamine, ×900). C: Transmission electron micrograph of area in B shows slowly advancing epithelium (E) to have produced only rare basement membrane (asterisk) and hemidesmosomes (arrowhead). A fibroblast (F) has infiltrated Bowman’s layer (×32,000). D: Phase-contrast photomicrograph at 10 weeks after a dilute sulfuric acid burn of a rhesus monkey cornea shows an irregular epithelial sheet separated (at asterisk) from the scarred Bowman’s layer and anterior stroma (paraphenylenediamine, ×600). E: Transmission electron micrograph of the corresponding area shows the epithelium to have become elevated with adherent fragments of basement membrane material (circled), allowing insinuation of flattened epithelial cells (E) with limited basement membrane deposition (asterisk; ×17,500). F: Light photomicrograph of the human cornea shown in A demonstrates characteristics of conjunctival overgrowth. Disorganized epithelium containing goblet cells (asterisks) overlies a thick fibrovascular pannus (P) that has developed anterior to the relatively intact Bowman’s layer (b; paraphenylenediamine, ×250). G: Transmission electron micrograph of an area shown in F shows that epithelial surface cells have normal microvillous surface membrane specialization (circled), and adjacent goblet cells have normal-appearing mucin globules (×18,700). |
Histologically, the conjunctival and corneal epithelia demonstrate surface keratinization with acanthosis and spongiosis. By transmission electron microscopy, keratohyalin granules are particularly visible in the thickened granular cell layer, as are increased desmosomes (93). Although actual bullous lesions are not seen clinically in the conjunctiva, ultrastructural examination discloses duplications of multilaminar basement membrane in the bulbar conjunctival epithelium, which may be related to the reformation of basement membrane complexes after resolution of an epithelial separation.
Mast cells are prominent participants in the conjunctival inflammatory reaction with varying degrees of vasculopathy or true vasculitis (92). These histologic and immunopathologic findings are similar to those in bullous pemphigoid, a nonocular bullous disease in which circulating antibodies react with a specific antigen localized to the lamina lucida portion of the BMZ (94). Because of these similarities, it may be appropriate to consider cicatricial mucous membrane pemphigoid and bullous pemphigoid as representing the same pathologic process. Recently, detailed histomorphometric analysis of human conjunctiva has demonstrated that tissue remodeling of the extracellular matrix is an essential and dynamic process that coexists with conjunctival scarring in ocular cicatricial pemphigoid and is linked with an immunoinflammatory process mediated by cytokines released by activated conjunctival cells and infiltrating cells; the final effect is a remodeling of the matrix in the conjunctival stroma, potentially regulating the altered metabolism of matrix proteins (95).
Persistent Defects and Recurrent Erosion of the Corneal Epithelium: Basement Membrane Disorders
As previously emphasized, the basement membrane complexes are responsible for the maintenance of tight adhesions between the corneal epithelium and stroma. The components of these complexes include hemidesmosomes of the basal epithelial cell membrane, the underlying basement membrane, and the subjacent anchoring fibrils of Bowman’s layer. Consequently, any traumatic, dystrophic, or degenerative disturbance at this level can predispose the epithelial cell layer to defective adhesion and repetitive breakdown, known as recurrent erosion syndrome (96). When the ocular surface is further compromised by extensive epithelial damage, inflammation, denervation, tear deficiency, or superficial stromal scarring, epithelial adhesion problems are compounded by a failure of epithelial mitosis or migration, or both, resulting in a persistent defect (96). In fact, when basement membrane is experimentally damaged by any technique, the consistent sequelae include irregularity and defects of the epithelial surface. Discontinuities and duplications of basement membrane complexes persist for 8 weeks or more, during which time the nonadherent epithelium is easily separable from the stroma (11,13). These same studies also emphasized the importance of the anterior stromal collagen of Bowman’s layer to which the basement membrane adheres. Although abnormalities of basement membrane complexes alone may explain poor adhesion and recurrent erosions in some situations, in others, damage to Bowman’s layer and anterior stroma with the accumulation of cellular and extracellular debris may also confound adhesion, despite the presence of basement membrane that appears morphologically normal (76).
Many clinical conditions that affect the corneal surface are manifested by recurrent erosions and persistent defects of the epithelium. These conditions can be considered as either primary or secondary causes of erosion, depending on whether the defect in the epithelial basement membrane is intrinsic or acquired. Among the primary disorders, epithelial basement membrane dystrophy (map-dot-fingerprint dystrophy, Cogan’s epithelial microcystic corneal dystrophy) is most common (Fig. 4-9A-G). This designation is based on the clinical morphologic finding of opaque intraepithelial cysts (dots), maplike geographic patterns, and whorled fingerprint-like ridges at the level of the epithelial basement membrane (97,98). Such changes have been noted in many people who are clinically asymptomatic, and in some families an apparent autosomal dominant inheritance has been determined (99). In symptomatic persons, the epithelium shows loose adhesion, and it may detach either spontaneously or after minor trauma. Thus, in patients presenting with recurrent erosion, slit-lamp examination of the uninvolved eye and of asymptomatic family members may establish an etiologic diagnosis.
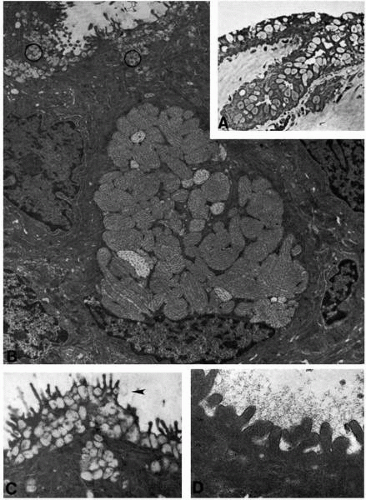 FIGURE 4-5. Human conjunctiva after chemical burn. A: Phase-contrast micrograph of conjunctival epithelium shows abnormally numerous light-stained goblet cells (paraphenylenediamine, ×250). B: Transmission electron microscopy shows a normal-appearing goblet cell in bulbar conjunctival epithelium, plus mucin granules (circles) in superficial epithelial cells (×8800). C: Higher magnification of mucin granules in nongoblet epithelial cells. Apparent secretion is indicated at the arrowhead (×18,700). D: At high magnification, epithelial microvilli with surface glycocalyx appear normal (×50,000). |
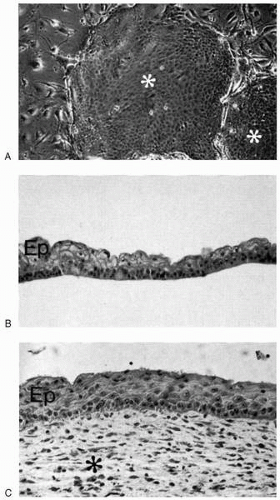 FIGURE 4-6. Xenograft transplantation of cultured human timbal cells. Phase-contrast microscopy (top) shows two colonies (asterisks) of cultured human limbal epithelial cells in culture surrounded by irradiated 3T3 fibroblasts (×100). Light microscopy (middle) of a confluent sheet of cultured limbal epithelial cells (Ep) displays considerable differentiation of compact basal cells and squamous superficial cells (hematoxylin-eosin, ×150). Light microscopy (bottom) of epithelium (Ep) formed from a sheet of cultured limbal epithelial cells grafted in the subdermal stroma (asterisk) of a nude mouse demonstrates cellular differentiation closely resembling corneal epithelium in vivo (hematoxylin-eosin, ×150). (Courtesy of Dr. Kristina Lindberg, with permission.) |
Recent application of in vivo confocal microscopy in patients with corneal recurrent erosion syndrome or epithelial basement membrane dystrophy showed deposits in basal epithelial cells, subbasal microfolds and streaks, damaged sub-basal nerves, or altered morphology of the anterior stroma, confirming clinically some observations achieved by other procedures (100). In fact, light and electron microscopic studies have also established that the microcystic areas correspond to the intraepithelial pseudocysts containing cellular debris. Geographic configurations represent thickened subepithelial plaques composed of abnormal multiple laminations of basement membrane and fibrillar collagens, whereas fingerprint lines consist of ridges of aberrant collagen and fibrous tissue (97,98,101). In these conditions, it is possible that dystrophic abnormalities of basement membrane complexes or adjacent structures permit the loosely adherent epithelium to shift, which would account for the accumulation of these abnormal configurations and for the predisposition to symptomatic erosions.
Although the effects can be variable, diabetes mellitus produces abnormalities of corneal epithelial adhesion. These corneal changes may also be considered a primary corneal basement membrane disorder. Ultrastructural studies of the human diabetic cornea have revealed an abnormality of the epithelial basement membrane that consists of thickening of the multilaminar basement membrane, duplication of anchoring fibrils, and decreased penetration of the anchoring fibrils from the deepest layer of the basal lamina into the stroma, which compromises the strength of the basement membrane complex at this level (96,102) (Fig. 4-9H-J). These changes are identical to those of the primary epithelial dystrophies. Although diabetic patients apparently do not experience an increased incidence of spontaneous epithelial erosions, vitreoretinal surgeons know that the diabetic corneal epithelium becomes clouded very early during surgery and often sloughs as an intact epithelial sheet. After surgery, these corneas are slow to reepithelialize, and persistent defects can develop in them (103). In nondiabetic humans and animals, corneal epithelial scraping results in rupture of basal cells with maintenance of basement membrane attached to Bowman’s layer. In the diabetic cornea, on the other hand, the entire epithelium separates as an intact sheet, with the entire basement membrane remaining adherent to the basal cells (96). This observation suggests that a major component of this erosive disorder is the lack of adhesion between the basement membrane complex and the underlying stroma caused primarily by decreased penetration of the anchoring fibrils into the stroma.
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


