Macular Oxygenation and Edema
Einar Stefánsson
INTRODUCTION
The supply of nutrition and oxygenation of the eye is unlike that for any other tissues in the body. In most other tissues, oxygen and nutrients are supplied to the cells from adjacent capillaries by diffusion over a concentration gradient that spans 50 to 100 micrometers at most. The eye on the other hand is mostly avascular, and cells must survive in areas where the nearest capillary is far away. The cornea receives oxygen in part from the atmosphere. The lens relies on nutrition and oxygen dissolved in the aqueous and vitreous humor. The cells in the vitreous humor also must rely on diffusion that carries oxygen and nutrients over almost a centimeter from the nearest capillaries in the retina.
It is obvious why the eye must be avascular. Visible light is absorbed by hemoglobin, and blood in the visual pathway impairs vision. A vascularized crystalline lens or vitreous humor (as in fetal stage) would render the eye virtually blind. The blood vessels in these tissues regress before birth and allow vision to develop unhindered. The visible light that we see must pass not only through the cornea, lens, and vitreous humor, but also through the retina itself. Fortunately, the retina is mostly transparent to visible light, with the notable exception of hemoglobin in the retinal blood vessels. While neural mechanisms mostly hide the presence of these vessels from our vision, we are able to visualize them in some conditions, such as when induced by blue light in the entoptic phenomenon.
The retinal circulation is in the pathway of light. In some species it is absent (guinea pig), partial and out of the way (rabbit), or minimal in extent (man). In the human, the retinal circulation only involves the inner retina, leaving the outer retina avascular and reliant on the choriocapillaris. The choriocapillaris supplies about 80% of the oxygen used by the retina. In addition, the inner retinal circulation is reduced to about 50% of the blood flow that a comparable volume of brain tissue would enjoy. The retina extracts more than 40% of the oxygen from the blood in the retinal circulation, whereas the average extraction for the brain and the body in general is 20% (Fig. 4-1). If the retina were vascularized in the same fashion as regular brain tissue, the amount of blood vessels and volume of blood would be about 10 times greater than it is. This would reduce the quality of vision considerably.
While the pattern of vascularization is very helpful for acute vision, it creates vulnerabilities. First, the inner retina normally has a very low oxygen tension, only about 20 mm Hg partial pressure, which is necessary for the extraction of 40% to 50% of the hemoglobin bound oxygen. It is operating on the edge in terms of oxygen tension and must employ a sophisticated autoregulation system to adapt to changes in oxygen supply or demand. Second, since the outer retina depends on oxygen and nutrients from the choriocapillaris, it is very sensitive to anything that disturbs or lengthens the diffusion path, including retinal detachment and subretinal hemorrhage.
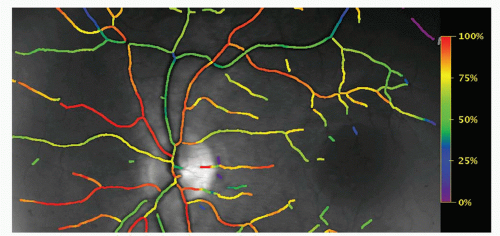 Figure 4-1. A color-coded map of hemoglobin oxygen saturation in the human retina. The map is generated automatically by a retinal oximeter. |
DOUBLE OXYGENATION
The retina receives oxygen from two sources, the retinal circulation and the choriocapillaris. The retinal circulation supplies the inner part of the retina and the choroid the avascular outer retina, including the photoreceptors. The blood flow in the retinal circulation is carefully regulated by a control mechanism known as “the autoregulation” and disturbances in inner retinal oxygenation generally result from occlusions of blood vessels, arterioles, capillaries or venules. On the other hand, disturbance in the oxygenation from the choroid are the result of an increased distance from the choriocapillaris to the retina or, less frequently, vascular occlusions in the choroid.
The blood flow in the choroid is relatively large. Whereas the retinal circulation loses about 40% to 50% of the available oxygen, the choroid only gives up about 4% of the available oxygen (1,2). This has led to some confusion about the role of the choroidal blood flow in the eye. Some have suggested that the main role of the choroid is to maintain temperature control in the fundus. While the choroid may contribute in this way, it is clear that the high blood flow is necessary to maintain a high enough oxygen tension in the choriocapillaris so that the oxygen tension gradient from the choriocapillaris to the mitochondria in the inner segments of the photoreceptors is steep enough to maintain the oxygen flux required. If the choroidal blood flow were so low that more hemoglobin dissociation would take place, the oxygen tension in the choroid and the oxygen flux to the photoreceptors would not meet the need of the rods and cones (Fig. 4-2).
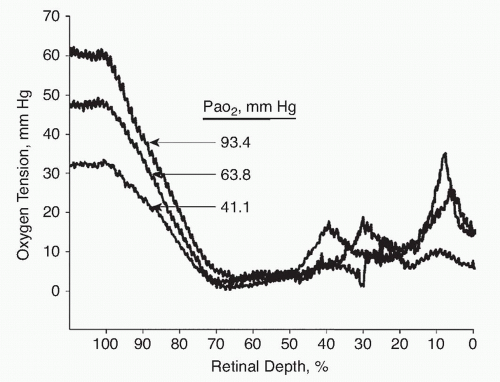 Figure 4-2. Intraretinal oxygen profiles recorded in dark during normoxia and hypoxemia. From The Journal of General Physiology 1992;99:177-197, with permission from The Rockefeller University Press. |
DISTURBANCE IN CHOROIDAL OXYGEN SUPPLY
Stefánsson (3) presented a mathematical model of the oxygenation of the retina. The model is based on Fick’s law, J = -D dC/dx, where J is the flux of oxygen, D the diffusion coefficient, and dC/dx the concentration gradient. If the distance between the retina and choriocapillaris is increased (dx is bigger), then the concentration gradient becomes flatter and the oxygen flux is decreased. Obviously, this takes place when a retinal detachment is present (3). Subretinal fluid in any form of retinal detachment, rhegmatogenous, exudative, or tractional, will decrease the oxygen flux from the choroid to the retina, and the same will be true if this distance is increased by pigment epithelial detachment, extensive drusen, or subretinal hemorrhage. Recent studies have shown the role of oxygen in retinal detachments (4).
AUTOREGULATION
Ocular blood flow follows the rule of Hagen-Poiseuille:
blood flow = perfusion pressure/vascular resistance F = (OAP-IOP)/R
The autoregulation influences vascular resistance (R) and thereby affects blood flow (F). OAP is the ophthalmic artery pressure and IOP the intraocular pressure. If metabolic requirements are constant, the autoregulation tries to keep blood flow constant. However, if metabolic requirements change, it changes the blood flow accordingly. The autoregulation does not primarily aim to maintain a constant rate of blood flow in the retina and optic nerve. It tries to maintain a constant chemical environment in the tissue. Blood flow is highly variable in normal retina. One example of this is that retinal blood flow varies with different O2 levels in breathing mixture (Fig. 4-3) (8).
Retinal blood flow also changes if the oxygen consumption of the retina is changed. Feke et al. (9) showed that human retinal blood flow is 40% to 70% lower in light than in dark. The change in retinal blood flow provides the appropriate change of available oxygen for the inner retinal tissue to compensate for increased oxygen consumption in the dark by the photoreceptor-RPE complex. The same group (10) showed that optic atrophy reduces retinal blood flow in humans. They found that blood flow in the temporal retinal arteries of the affected eyes, measured by the laser Doppler technique, was 48% +/− 20% lower than in the fellow eyes (four patients). Arterio-venous O2 saturation difference for temporal and nasal vascular segments of the affected eyes, evaluated by retinal vessel oximetry, was 12% +/− 9% higher than in the fellow eyes. The combination of these results indicated a 40% +/− 29% reduction in O2 delivery in the affected eyes, thereby quantifying the decrease in retinal metabolism that resulted from inner retinal degeneration.
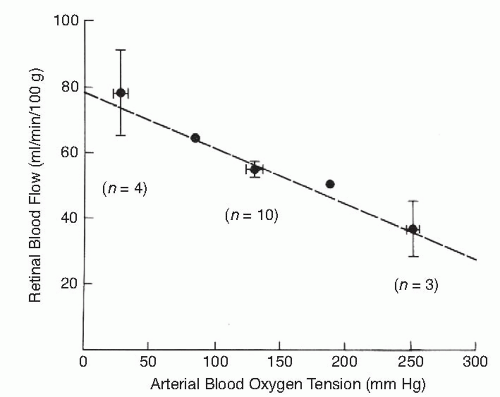 Figure 4-3. The relationship of retinal blood flow with arterial blood oxygen tension, measured in cats. (From Stefansson E, Wagner HG, Seida M. Retinal blood flow and its autoregulation measured by intraocular hydrogen clearance. Exp Eye Res 1988;47:669-678, with permission). |
Naturally, if the metabolic demand is constant, the retinal autoregulation will try to keep blood flow and the chemical environment constant. If the IOP is changed (and the OAP is constant), the perfusion pressure will change. In this case the autoregulation will dilate the arterioles, decrease the resistance (R) and maintain constant blood flow. Figure 4-4 (11,12) shows the oxygen tension over the optic nerve in pigs, where the IOP is stepwise changed. With moderate changes in IOP the oxygen tension does not change, as the autoregulation keeps it constant. However, a large change in IOP overwhelms the autoregulation and oxygen tension falls.
STARLING’S LAW AND MACULAR EDEMA
Ernest Henry Starling (1866, 1867, 1868, 1869, 1870, 1871, 1872, 1873, 1874, 1875, 1876, 1877, 1878, 1879, 1880, 1881, 1882, 1883, 1884, 1885, 1886, 1887, 1888, 1889, 1890, 1891, 1892, 1893, 1894, 1895, 1896, 1897, 1898, 1899, 1900, 1901, 1902, 1903, 1904, 1905, 1906, 1907, 1908, 1909, 1910, 1911, 1912, 1913, 1914, 1915, 1916, 1917, 1918, 1919, 1920, 1921, 1922, 1923, 1924, 1925, 1926 and 1927) stated in 1896: “There must be a balance between the hydrostatic pressure of the blood in the capillaries and the osmotic attraction of the blood for the surrounding fluids…. and whereas capillary pressure determines transudation, the osmotic pressure of the proteins of the serum determines absorption.” In other words, the hydrostatic pressure forcing fluids from the vessel into the tissue must be balanced by the osmotic pressure, generated by the colloidal protein solutions in the capillary, forcing absorption of the fluid from the tissues (13).
Starling’s four forces that govern the transport of water between the vascular compartment and the tissue compartment are the following: (a) Hydrostatic pressure in the capillary (Pc). (b) Hydrostatic pressure in the interstitium (Pi), which in the eye equals the intraocular pressure, IOP. (c) Osmotic (oncotic) pressure exerted by plasma proteins in the capillary (Qc). (d) Osmotic pressure exerted by proteins in the interstitial fluid (Qi).
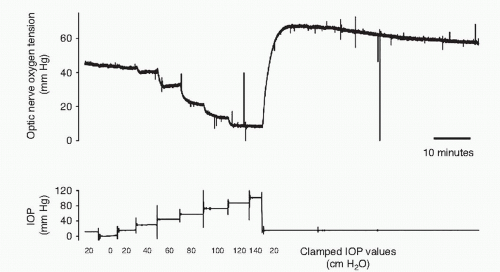 Figure 4-4. Optic nerve oxygen tension measured with a polarographic oxygen electrode placed in the vitreous approximately 0.5 mm above the optic nerve head in a pig. The intraocular pressure (IOP) was controlled by means of a canula placed in the anterior chamber and connected to a saline reservoir. Intraocular pressure was continuously regulated and optic nerve oxygen tension measured continuously at the same time. (From la Cour M, Kiilgaard JF, Eysteinsson T, et al. Optic nerve oxygen tension: effects of intraocular pressure and dorzolamide. Br J Ophthalmol 2000;84:1045-1049, with permission.) |
The balance of these forces allows the calculation of the net driving pressure for filtration
Net Driving Pressure = (Pc − Pi) − (Qc − Qi)
The hydrostatic pressure, which originates in the heart, is higher in the vessel than in the tissue and this drives water from the vessel into the tissue. The hydrostatic pressure gradient, δP, must be balanced by the osmotic pressure gradient, δQ, where the osmotic pressure is higher in the blood than in the interstitial fluid and pulls water back into the blood vessel. If the hydrostatic pressure gradient and the osmotic pressure gradient are equal, no net transport of water takes place and edema is neither formed nor resolved. Starling’s law is frequently shown in this form as
δP − δQ = 0
describing the steady state of the equal and opposing hydrostatic, δP, and osmotic pressure, δQ, gradients (13).
Starling’s law has been generally accepted in medicine and physiology for more than a century as the fundamental rule governing the formation and disappearance of edema in the body. It is reasonable to believe that the ocular tissues follow the same general laws of physiology and physics as the rest of the body, and those who believe otherwise should be burdened with the duty of disproving Starling’s law in the eye.
EDEMA
Edema is the swelling of soft tissues due to an abnormal accumulation of fluid, that is, water. Edema may be cytotoxic or vasogenic in origin. In cytotoxic or ischemic edema the abnormal water accumulation and swelling occurs within cells (14), whereas in vasogenic edema the water accumulates in the interstitial space between cells. While retinal edema may be either cytotoxic or vasogenic, Starling’s law applies to the vasogenic edema, which presumably is the most frequent and important form of edema in vascular retinopathies.
With abnormal accumulation of water in the retina the tissue volume increases and the retina thickens. The thickening may be measured with ocular coherent tomography (OCT). At the same time the specific gravity of the tissue is decreased proportionally with the increased water content (15).
WHAT CREATES EDEMA?
According to Starling’s law, edema will form if the hydrostatic pressure gradient between vessel and tissue is increased or the osmotic pressure gradient is decreased. The hydrostatic gradient increases if the blood pressure in the microcirculation rises or the tissue pressure decreases. The osmotic pressure gradient decreases if proteins accumulate in the interstitium to increase the osmotic pressure in the tissue, and also if the osmotic pressure in blood goes down.
Increased Hydrostatic Pressure Gradient
The hydrostatic pressure in the microcirculation, capillaries, and venules is a function of the work of the heart, arterial blood pressure, and the resistance and pressure fall in the arterioles. Arterial hypertension tends to increase the hydrostatic pressure in the capillaries and is a well-known risk factor for diabetic macular edema (16, 17, 18, 19, 20 and 21). Diabetic macular edema tends to improve if arterial hypertension is successfully treated (17,18).
The resistance in the retinal arterioles and thereby the pressure drop in the arterioles is a function of the diameter of
the arterioles. The resistance to flow is described by the Hagen-Poiseuille law, where the resistance is inversely related to the fourth power of the vessel radius (13). If the arterioles dilate, as they do in hypoxia, the resistance in the arterioles decreases and the hydrostatic pressure in the capillary bed rises (22, 23 and 24



the arterioles. The resistance to flow is described by the Hagen-Poiseuille law, where the resistance is inversely related to the fourth power of the vessel radius (13). If the arterioles dilate, as they do in hypoxia, the resistance in the arterioles decreases and the hydrostatic pressure in the capillary bed rises (22, 23 and 24
Stay updated, free articles. Join our Telegram channel
Full access? Get Clinical Tree
