(Fig. 27-2) (6,7). The goblet cells of the conjunctiva and the stratified squamous cells of the cornea and conjunctiva secrete the mucous layer of the tear film (Fig. 27-2). These stratified squamous cells also produce the glycocalyx. The lacrimal gland also secretes a soluble mucin.
TABLE 27-1. PHYSICAL CHARACTERISTICS OF TEARS | ||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ||||||||||||||||||
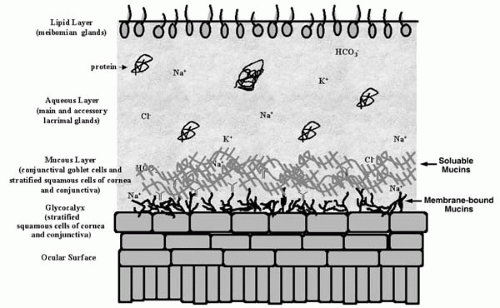 FIGURE 27-1. Schematic drawing of the three layers of the tear film. The tear film covers the cells of the ocular surface (cornea and conjunctiva). The upper lipid layer is secreted by the meibomian glands; the middle aqueous layer is secreted by the main and accessory lacrimal glands and the conjunctival epithelium; the inner mucous layer is secreted by conjunctival goblet cells and stratified squamous cells of the conjunctiva and cornea and the glycocalyx of the cornea and conjunctiva. (From Hodges and Dartt. Regulatory pathways in lacrimal gland epitheliom. Int Rev Cytol 2003;231:129-196.) |
a plethora of lipids, which melts at about 35°C and thus is always fluid in the living eye. The lipid layer behaves as a film essentially independent of the aqueous layer underneath. It is anchored at the orifices of the meibomian glands above and below and does not take part in the flow of tears from the lateral canthus to the lacrimal puncta (1). When the lids close during a blink, the lipid layer is compressed over the aqueous layer (14). When the lids open, the lipid layer begins to spread again over the aqueous layer. The spreading front can move faster than the opening lid, so the aqueous layer is never exposed (14). During lid closure the lipid layer thickens, and during eye opening it thins. This has been elegantly shown by Doane (15) using a specially designed interferometer that detects colors produced by interference patterns as the light passes through an oily layer whose thickness is changing.
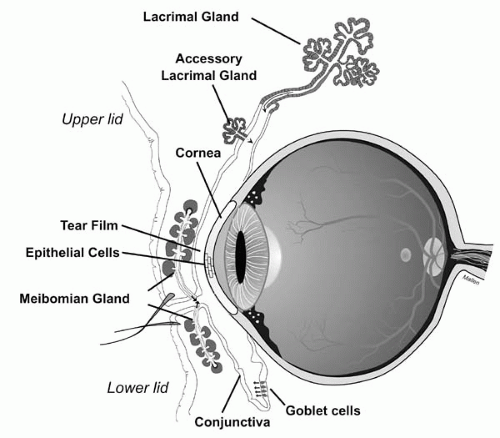 FIGURE 27-2. Schematic representation of the tear-producing glands. Cross section of the eye and the glands that produce the three layers of the tear film. (From Dartt DA. Regulation of mucin and fluid secretion by conjunctival epithelial cells. Prog Retin Eye Res 2002;21:555-576.) |
This accounts for the complex nature of the meibomian gland fluid. This fluid is stored in the ducts until released by the action of the blink.
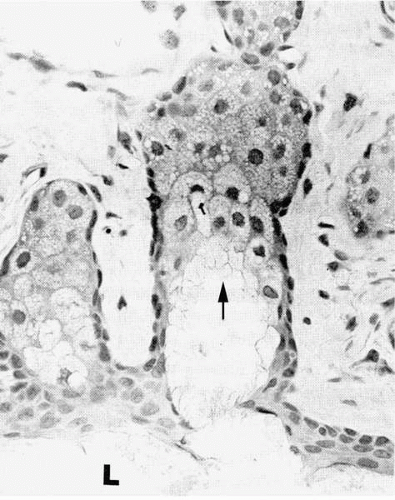 FIGURE 27-3. Light micrograph of alveoli of a rabbit meibomian gland. Cells in the outer layer (arrow) of the alveolus do not contain lipid droplets. Clear lipid droplets are visible in the middle layers of cells. Cells in the inner layer disintegrate into the alveolar duct, releasing the lipid droplets to form the secretory product that diffuses into the lumen (L) of the central duct. (H&E ×350.) (From Jester JV, Nicolaides N, Smith RE. Meibomian gland dysfunction. I: Keratin protein expression in normal human and rabbit meibomian glands. Invest Ophthalmol Vis Sci 1989;30:927.) |
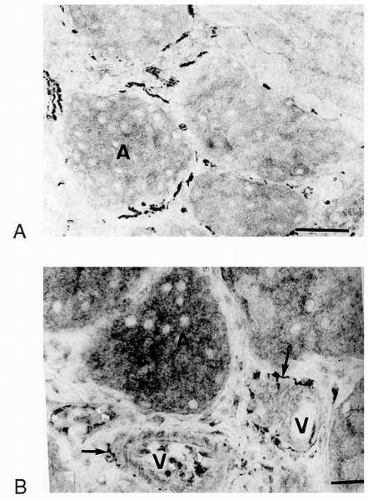 FIGURE 27-4. Innervation of the rat meibomian glands. Immunohistochemical microscopy showing localization of (A) sympathetic nerves using an antibody against tyrosine hydroxylase, which is one of the enzymes that coverts dopamine to norepinephrine, and (B) parasympathetic nerves using an antibody against choline acetyltransferase. (From Seifert P, Spitznas M. Immunocytochemical and ultrastructural evaluation of the distribution of nervous tissue and neuropeptides in the meibomian gland. Graefes Arch Clin Exp Ophthalmol 1996;234:648-656.) |
sterol esters, free fatty acids, and polar lipids (for a review see ref. 26). The complexity of meibomian gland fluid reflects the products from the disintegrating cells as well as from the synthesized lipids. Meibomian gland lipids differ from cellular lipids in the acyl chain types and the sterol types. The major classes of lipids are the wax monoesters and sterol esters that make up about 60% to 70% of the meibomian gland fluid (26). A third type of lipid is the diesters that form ester linkages with fatty acids, fatty alcohols, or sterols. These diester compounds make up about 8% of the fluid.
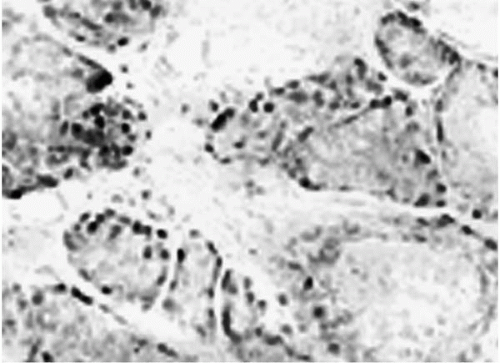 FIGURE 27-5. Presence of androgen receptor protein in acinar cells of the rat meibomian gland. Immunoperoxidase microscopy of meibomian gland showing the location of androgen receptor protein in acinar cells. (From Sullivan DA, Sullivan BD, Ullman MD, et al. Androgen influence on the Meibomian gland. Invest Ophthalmol Vis Sci 2000;41:3732-3742.)(see color image) |
ductal cells to eject the secretory product (27). Many types of lymphocytes producing immunoglobulin (Ig) A, IgG, IgE, IgM, and IgD, plasma cells, mast cells, and macrophages are also present in the lacrimal gland.
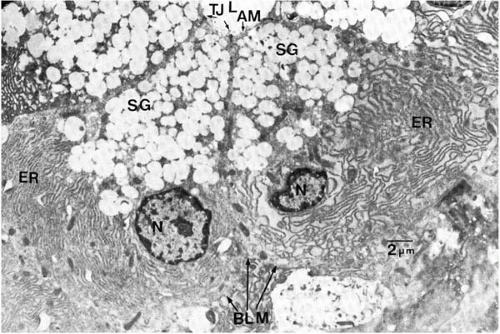 FIGURE 27-6. Electron micrograph of adjacent acinar cells from the rat lacrimal gland. The nucleus (N) lies in the basal portion of the cell and is surrounded by endoplasmic reticulum (ER). Secretory granules (SG) lie in the apical portion of the cell. Tight junctions (TJs) are present adjacent to the lumen (L) separate the apical membrane (AM) from the basolateral membrane (BLM). (From Dartt DA. Signal transduction and control of lacrimal gland protein secretion: a review. Curr Eye Res 1989;8:619-636.) |
TABLE 27-2. PROTEINS KNOWN TO BE SECRETED BY THE LACRIMAL GLAND | ||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
|
is complicated due to the fact that 11 different isoforms of PKC have been identified. The lacrimal gland contains PKC-α, δ, ε, and λ. In general, PKC isoforms have cell and tissue specific localizations, and the lacrimal gland is no exception (28). The PKC isoforms detected in the lacrimal gland have specific although somewhat overlapping distribution (28).
TABLE 27-3. GENDER-RELATED DIFFERENCES IN THE LACRIMAL GLAND AND TEAR FILM | |||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| |||||||||||||||||||||||||||||||||||||||
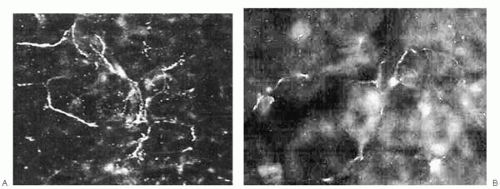 FIGURE 27-7. Innervation of the rat lacrimal gland. Immunofluorescent microscopy showing localization of (A) parasympathetic nerves using an antibody against vasoactive intestinal peptide (VIP). B. Sympathetic nerves using an antibody against dopamine β-hydroxylase which is the enzyme that coverts dopamine to norepinephrine. |
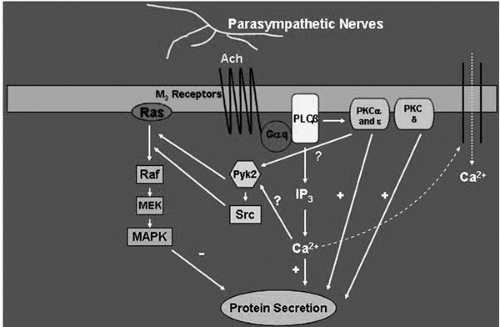 FIGURE 27-8. Schematic drawing of the signal transduction pathways activated by cholinergic agonists in the lacrimal gland. Ach, acetylcholine; M3, muscarinic receptor subtype 3; Gαq, alpha subunit of Gq G protein; PLC, phospholipase C; Pyk2 and Src, nonreceptor tyrosine kinases; raf, mitogen-activated protein kinase kinase kinase; MEK, mitogen-activated protein kinase kinase; MAPK, p44/p42 mitogen-activated protein kinase; InsP3, inositol trisphosphate; PKC, protein kinase C. (Modified from Dartt DA. Regulation of lacrimal gland secretion by neurotransmitters and the EGF family of growth factors. Exp Eye Res 2001;73:741-752.) |
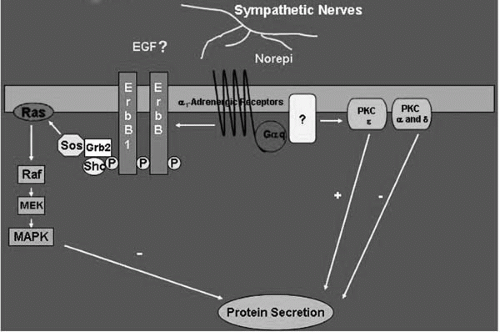 FIGURE 27-9. Schematic drawing of the signal transduction pathways activated by α1-adrenergic agonists in the lacrimal gland. Norepi, norepinephrine; PKC, protein kinase C; ErbB1, the epidermal growth factor (EGF) receptor; ErbB, the EGF family of growth factor receptor; P, phosphorylated tyrosine residues on the ErbBs; SOS, Grb2 and Shc, adaptor proteins; raf, mitogen-activated protein kinase kinase kinase; MEK, mitogen-activated protein kinase kinase; MAPK, p44/p42 mitogen-activated protein kinase. (Modified from Dartt DA. Regulation of lacrimal gland secretion by neurotransmitters and the EGF family of growth factors. Exp Eye Res 2001;73:741-752.)
Stay updated, free articles. Join our Telegram channel
Full access? Get Clinical Tree
 Get Clinical Tree app for offline access
Get Clinical Tree app for offline access

|