Past theories that have been proposed to account for the attacks of vertigo during the course of Meniere disease are reviewed. In the past, vascular theories and theories of perilymph and endolymph mixing due to ruptures or leakages were proposed. Recent research concerning the basic mechanisms of the inner ear anatomy and function cast doubt on these theories. The anatomy, physiology, and pathophysiology of the inner ear, and in particular of the endolymphatic sac and endolymphatic duct are reviewed. Recent studies suggest that in people the endolymph ionic content is replenished without any flow of fluid and that longitudinal endolymph flow only occurs in response to volume excess. Furthermore audiological and electrophysiological studies have revealed little or no change in the cochlear function during episodes of vertigo. The longitudinal drainage theory attempts to encompass the recent research findings. The theory hypothesizes that endolymph draining too rapidly from the cochlear duct (pars inferior) causes attacks of vertigo. The endolymph overfills the endolymphatic sinus and overflows into the utricle (pars superior), stretching the cristae of the semicircular canals, causing the attacks of vertigo.
Vertigo is the major symptom of Meniere’s disease during the early stages of the disorder. The vertigo can be very unpleasant and disabling and can occur with little warning. Characteristically there is a sensation of movement, usually a feeling of rotation, associated with nausea and vomiting. Each attack occurs for least 10 minutes and may last for several hours. Each attack is separated from the next by at least one day. The attacks of vertigo tend to occur in clusters over a period of several weeks followed by variable periods of remission. Sometimes the periods of remission may last for several years, or the disorder may completely resolve.
Understanding the pathophysiological mechanism that causes the attacks of vertigo could provide the key to understanding the etiology of Meniere’s disease.
Before Prosper Ménière’s classic description of the disease, it was thought the vertigo attacks were caused by cerebral apoplexy or a type of epilepsy. Ménière correctly attributed the attacks to a disorder of the inner ear, but his original paper was met with scorn and skepticism. Ménière suggested the mechanism could be similar to migraine, and for a while the popular theory was that vasospasm within the inner ear caused the attacks. Different therapies were based on this hypothesis, including cervical sympathectomy and vasodilator medications such as nicotinic acid.
In 1938, Hallpike and Cairns in London and Kyoshiro Yamakawa in Japan both independently published temporal bone histologic studies that showed the presence of endolymphatic hydrops. Endolymphatic hydrops is a term to describe increased volume of endolymph within the membranous inner ear. It also was commonly supposed that there was an increased pressure of endolymph causing the attacks.
In 1964, Schuknecht proposed the rupture theory, based on his histologic studies, where he identified areas of Reissner membrane that showed evidence of healing after ruptures. He suggested that these might have occurred because of increased endolymph volume and that the ruptures in Reissner membrane led to mixing of potassium-rich endolymph with perilymph. The afferent vestibular nerves were paralyzed by the high potassium until the ionic pumps within the inner ear restored the electrolyte levels and the rupture healed.
The rupture theory remained the most plausible explanation for several decades. The theory led to the concept that it was blockage of flow along the endolymphatic duct that led to endolymphatic hydrops. Endolymphatic sac surgery was believed to unblock the duct and allow the endolymphatic sac to function effectively again, clearing the excess endolymph and restoring normal inner ear function.
As knowledge of the physiology and pathophysiology of the ear has developed over recent years, the original rupture theory hypothesis has seemed increasingly unlikely, and alternative theories have evolved to explain a sudden potassium contamination of the perilymph. For example, leakage of potassium through gaps in the tight junctions and changes in the calcium levels controlling tight junctions in vestibular hair cells have been suggested. But is the vertigo truly a result of potassium leakage, or should an alternative hypothesis be sought? The anatomy and physiology of the inner ear are complex. Great strides in understanding have occurred, however, and this must be considered before formulating any hypothesis ( Fig. 1 ).
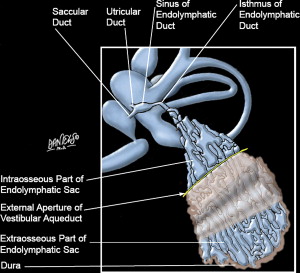
The anatomy of the endolymphatic sac and duct
The anatomy of the human endolymphatic duct and sac is shown in Fig. 1 . The duct begins at the ductus reuniens, which joins the cochlear duct and the utricle to form an endolymphatic duct (ED) leading to the endolymphatic sac (ELS). The anatomy has been described by Lo and his colleagues. Often anatomic texts describe a long thin ED ending in a short, pouch-like ELS, but in reality, the system is far different in appearance. The ED is a short single-lumen tube of only 2 mm in length. The ELS is much larger and a highly complex structure of interconnecting tubules, cisterns and crypts. The endolymphatic sinus (ES) lies in a groove on the posteromedial surface of the vestibule, with its distal outlet leading to the vestibular aqueduct. The ED narrows at its isthmus at the isthmus of the vestibular aqueduct (VA), where it is oblong in shape with mean diameter of only 0.09 × 0.2 mm. Distal to the isthmus begins the ELS, which flares considerably transversely but thickens only slightly in its sagittal dimension. The ELS has two portions, the intraosseous portion within the VA, and the extraosseous portion. The size of the intraosseous portion varies considerably, from 6 to 15 mm in length and 3 to 15 mm in width. The extraosseous ELS rests on a fovea on the posterior wall of the petrous bone, where the ELS lies between two layers of dura. The extraosseous ELS also varies considerably in size, from 5 to 7 mm in width and 10 to 15 mm in length.
In lower animals and the human fetus, the ELS consists of a single lumen. In people after the age of 1 year, the ELS develops tubules that reach adult complexity by the age of 3 to 4 years. The tubules of the ELS are more complex in the proximal and middle (rugose) portions. The concept of the human ELS as a empty sack is entirely wrong.
The bony (VA) has a smaller diameter in Meniere’s ears than the median in normal ears. Basically all Meniere’s ears have a small VA, but there are non-Meniere’s ears that also have small-diameter VA.
The physiology of the endolymphatic sac and its role in longitudinal endolymph flow
The ELS has absorptive and secretory functions as well as phagocytic and immune defence functions. It has been known for many years that debris placed into the cochlear duct passes to the ELS. How the debris reaches the endolymphatic sac has been a matter of conjecture but there must be longitudinal flow of endolymph.
The endolymphatic sac secretes various substances, including aquaporins, glycoproteins, and even endolymph. Glycoprotein is highly hydrophilic, and studies suggest that when longitudinal flow occurs, the glycoprotein is rapidly eaten away by phagocytic activity. It is possible that the glycoprotein is both produced and eaten away to cause longitudinal flow. Magnetic resonance imaging (MRI) studies suggest activity occurs in the endolymphatic sac during attacks of Meniere’s disease.
Salt has undertaken some very important work on how endolymph is produced and how it circulates. He has shown that in guinea pigs, the composition of endolymph is maintained by the stria vascularis. The stria vascularis controls the influx of water and modifies the ionic content. Normally there is little flow either radially or longitudinally, and the endolymph is a biologic puddle. If excess endolymph volume occurs, this is reabsorbed back into the stria vascularis (radial flow). Only under exceptional circumstances, such as when there is a large volume increase, does the endolymph move longitudinally to the endolymphatic sac. People can maintain the balance of endolymph by the radial mechanism alone and rarely have to call upon the endolymphatic sac mechanisms to provide longitudinal flow.
The rate of longitudinal flow is restricted by the isthmus of the ED rather like sand draining through an hour glass. It is hypothesized that if the endolymph cannot drain quickly enough, the endolymphatic sinus may act as a reservoir, temporarily holding the excess fluid.
The physiology of the endolymphatic sac and its role in longitudinal endolymph flow
The ELS has absorptive and secretory functions as well as phagocytic and immune defence functions. It has been known for many years that debris placed into the cochlear duct passes to the ELS. How the debris reaches the endolymphatic sac has been a matter of conjecture but there must be longitudinal flow of endolymph.
The endolymphatic sac secretes various substances, including aquaporins, glycoproteins, and even endolymph. Glycoprotein is highly hydrophilic, and studies suggest that when longitudinal flow occurs, the glycoprotein is rapidly eaten away by phagocytic activity. It is possible that the glycoprotein is both produced and eaten away to cause longitudinal flow. Magnetic resonance imaging (MRI) studies suggest activity occurs in the endolymphatic sac during attacks of Meniere’s disease.
Salt has undertaken some very important work on how endolymph is produced and how it circulates. He has shown that in guinea pigs, the composition of endolymph is maintained by the stria vascularis. The stria vascularis controls the influx of water and modifies the ionic content. Normally there is little flow either radially or longitudinally, and the endolymph is a biologic puddle. If excess endolymph volume occurs, this is reabsorbed back into the stria vascularis (radial flow). Only under exceptional circumstances, such as when there is a large volume increase, does the endolymph move longitudinally to the endolymphatic sac. People can maintain the balance of endolymph by the radial mechanism alone and rarely have to call upon the endolymphatic sac mechanisms to provide longitudinal flow.
The rate of longitudinal flow is restricted by the isthmus of the ED rather like sand draining through an hour glass. It is hypothesized that if the endolymph cannot drain quickly enough, the endolymphatic sinus may act as a reservoir, temporarily holding the excess fluid.
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


