TABLE 3-1. CLUSTERS OF DIFFERENTIATION (CD) DESIGNATIONS | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
subtypes occurs in the spleen. Macrophages that express a high density of class II MHC glycoproteins are present in red pulp, and macrophages with significantly less surface class II MHC glycoprotein expression are in the marginal zone, where intimate contact with B cells exists. It is likely that, just as in the murine system (10), so too in humans, one subclass of macrophage preferentially presents antigen to one particular subset of helper T cells responsible for induction of regulatory T-cell activation, whereas a different subset of macrophage preferentially presents antigen to a different helper T-cell subset responsible for cytotoxic or delayed-type hypersensitivity effector functions.
TABLE 3-2. NEUTROPHIL GRANULES AND THEIR CONTENTS | |||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
|
TABLE 3-3. GRANULAR CONTENT OF EOSINOPHILS | ||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
|
TABLE 3-4. MAST CELL TYPES AND CHARACTERISTICS | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
the blood islands of the yolk sac (13), and populate embryonic liver and bone marrow (14). These stem cells are pluriopotential. Characteristics of the microenvironment in the bone marrow, particularly with respect to a stem cell’s association with other resident cells in the bone marrow, contribute to or are responsible for the different pathways of maturation and differentiation. For example, specific cells in the bone marrow in the endosteal region promote the differentiation of hematopoietic stem cells into B lymphocytes (15). In birds, primordial pluripotential stem cells that migrate to a gland near the cloaca of the chicken known as the bursa of Fabricius (for reasons of probable stimuli in the bone marrow not yet understood) are influenced by the epithelial cells in that gland to differentiate terminally into B lymphocytes (16,17).
TABLE 3-5. MAST CELL CONTENTS | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
|
TABLE 3-6. PLATELET GRANULES AND THEIR CONTENTS | ||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ||||||||||||||||||||||||||||||||||||||||
receptors on the surface of mature T cells have been identified for the lymph node (MEL-14 or L-selectin [leukocyte functional antigen (LFA)-1]) and for Peyer’s patch [leukocyte Peyer’s patch adhesion molecule (LPAM)-1 α4β7 integrin, CD44]. Equivalent homing receptors undoubtedly exist in humans, but work in this area is currently embryonic. A 90-kD glycoprotein designated Hermes-3, however, has been identified as a specific heterotypic recognition unit on lymphocytes (20). The Hermes glycoprotein has been shown to be identical to the CD44 molecule (21). Antibodies to this glycoprotein prevent binding of lymphocytes to mucosal lymph node high-endothelial venules (22,23).
TABLE 3-7. LYMPHOID ORGANS | ||||||||
|---|---|---|---|---|---|---|---|---|
|
cytokine genes is then activated and regulated by it (them). For example, NFATc translocates to the nucleus, where it combines with adaptor proteins (AP-1); this complex then binds to the NFAT-binding site of the IL-2 promoter. This, coupled with nuclear factor-kappa B binding by proteins possibly induced by the events stimulated by CD28-CD80 signal transduction, results in IL-2 gene transcription typical of T-cell activation (Fig. 3-1). Thus, this activation phase of the acquired immune response is characterized by lymphocyte proliferation and cytokine production.
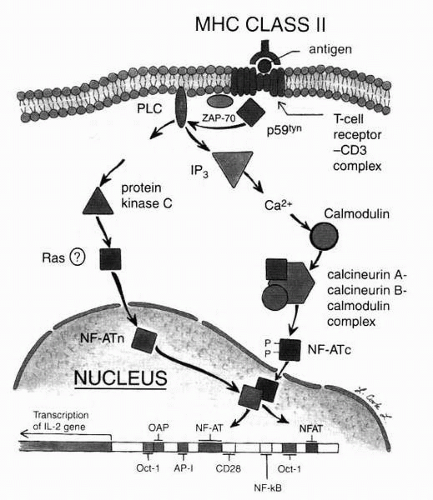 FIG. 3-1. Signal transduction: intracellular and intranuclear. With antigen-presenting cell presentation of antigen to the T cell [green peptide fragment in the major histocompatibility complex (MHC) class II groove of the macrophage], an extraordinary cascade of events occurs, through the cell membrane, into the cytoplasm, and subsequently into the nucleus, to the level of specific genes on the chromosomes of the nucleus. Specifically, tyrosine-rich phosphorylases catalyze phosphorylation of a series of intracellular proteins, with resultant liberation of calcium stores, and production of the calcineurin-calmodulin complex, which then facilitates the production of nuclear factor of activated T cells, cytoplasmic component capable of being transported through one of the nuclear pores into the nucleus, where interaction then with specific foci on the gene results in induction of gene transcription (in this instance, production of messenger RNA for ultimate synthesis of the protein interleukin-2). (Original drawing by Laurel Cook Lowe.) |
TABLE 3-8. ADHESION MOLECULES | |||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| |||||||||||||||||||||||||||||||||||||
number of lymphocytes derived from the original clone of cells stimulated by primary contact with antigen, and because of the longevity of many of the cells (memory cells). The memory cells can survive for very long periods, even decades. They express cell surface proteins not expressed by nonmemory cells (CD45RO). In memory cells, the level of cell surface expression of peripheral lymph node homing receptors is low compared with the population of such receptors on the surfaces of nonmemory cells; in contrast, the population of other adhesion molecules includes CD11a, CD18 (LFA-1), CD44, and HLA molecules. Because of the constitutive expression of the cell surface adhesion molecules, memory T cells rapidly home to sites of inflammation, “looking” for antigen to which they might respond.
TABLE 3-9. CYTOKINES AND TARGET CELLS | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
possibility that immunoglobulin genes possess an unusual architecture. In 1965, Dreyer and Bennett proposed that the V and C regions of an immunoglobulin chain are encoded by two separated genes in embryonic (germline) cells (germline gene diversity) (24). According to this model, one of several V genes becomes joined to the C gene during lymphocyte development. In 1976, Hozumi and Tonegawa discovered that V and C regions are encoded by separate, multiple genes far apart in germline DNA that become joined to form a complete immunoglobulin gene active in B lymphocytes (25). Immunoglobulin genes are thus translocated during the differentiation of antibody-producing cells (somatic recombination; Fig. 3-3).
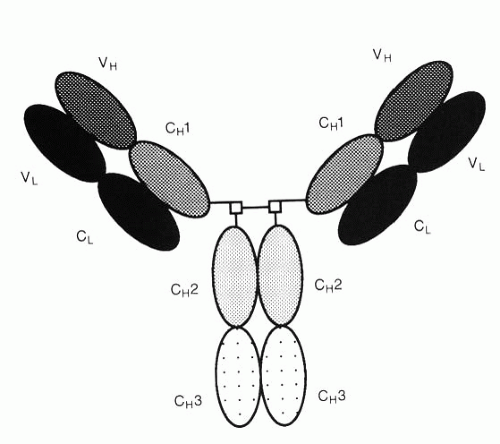 FIG. 3-2. Structure of immunoglobulin G showing the regions of similar sequence (domains). (From Albert DA, Jakobiec FA. Principles and practice of ophthalmology, 2nd ed. Philadelphia: WB Saunders, 2000:66.) |
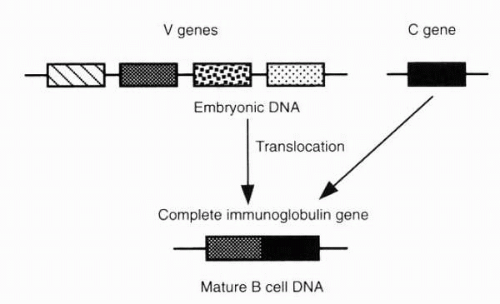 FIG. 3-3. Translocation of a V-segment gene to a C gene in the differentiation of an antibody-producing B cell. (From Albert DA, Jakobiec FA. Principles and practice of ophthalmology, 2nd ed. Philadelphia: WB Saunders, 2000:67.) |
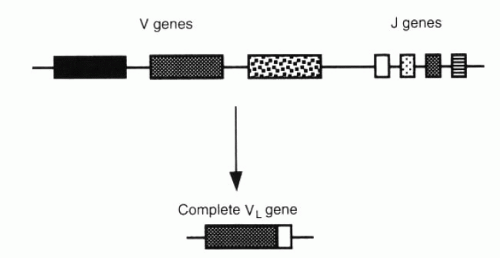 FIG. 3-4. Hypervariable or complementarity-determining regions (CDRs) on the antigen-binding site of the variable regions of immunoglobulin G. (From Albert DA, Jakobiec FA. Principles and practice of ophthalmology, 2nd ed. Philadelphia: WB Saunders, 2000:67.) |
 FIG. 3-5. A V gene is translocated near a J gene in forming a light-chain V region gene. (From Albert DA, Jakobiec FA. Principles and practice of ophthalmology, 2nd ed. Philadelphia: WB Saunders, 2000:67.) |
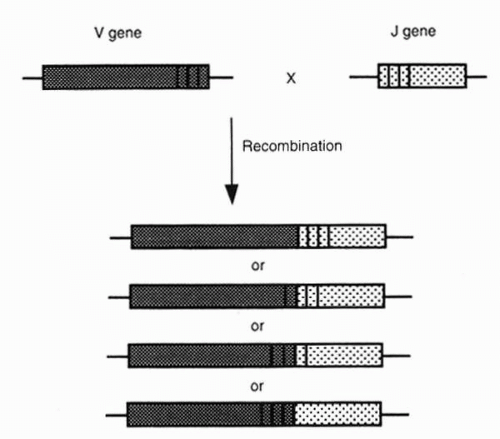 FIG. 3-6. Imprecision in the site of splicing of a V gene to a J gene (junctional diversity). (From Albert DA, Jakobiec FA. Principles and practice of ophthalmology, 2nd ed. Philadelphia: WB Saunders, 2000:68.) |
be generated by combining different V, D, and J gene segments and by combining more than 3000 L chains and 45,000 H chains. If the effects of N-region additional are included, more than 1011 different combinations can be formed. This is large enough to account for the immense range of antibodies that can be synthesized by an individual.
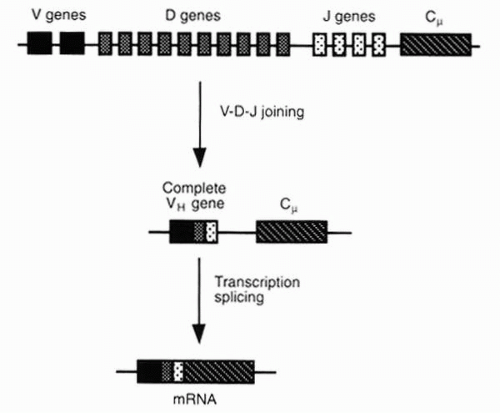 FIG. 3-7. The variable region of the heavy chain is encoded by V-, D-, and J-segment genes. (From Albert DA, Jakobiec FA. Principles and practice of ophthalmology, 2nd ed. Philadelphia: WB Saunders, 2000:68.) |
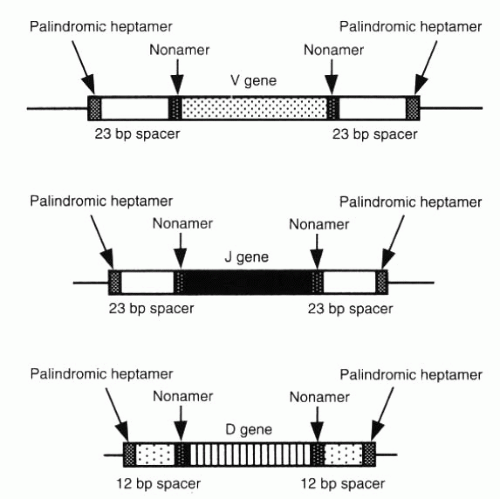 FIG. 3-8. Recognition sites for the recombination of V-, D-, and J-segment genes. V and J genes are flanked by sites containing 23-bp spacers, whereas D-segment genes possess 12-bp spacers. Recombination can occur only between sites with different classes of spacers. (From Albert DA, Jakobiec FA. Principles and practice of ophthalmology, 2nd ed. Philadelphia: WB Saunders, 2000:68.) |
cytokines produced by helper T cells (28). For example, switching to IgE class immunoglobulin production is provoked by the CD4 TH2 cytokine, IL-4.
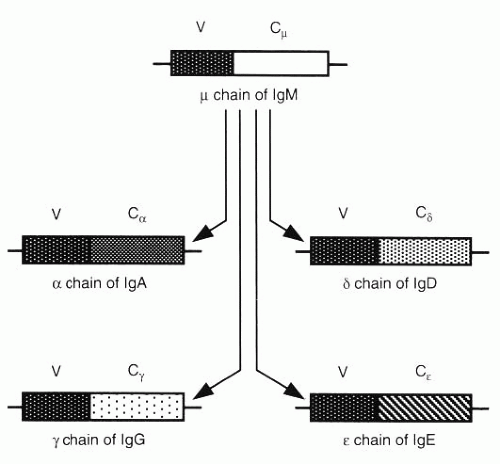 FIG. 3-9. The VH region is first associated with Cμ and then with another C region to form an H chain of a different class in the synthesis of different classes of immunoglobulins. (From Albert DA, Jakobiec FA. Principles and practice of ophthalmology, 2nd ed. Philadelphia: WB Saunders, 2000:69.) |
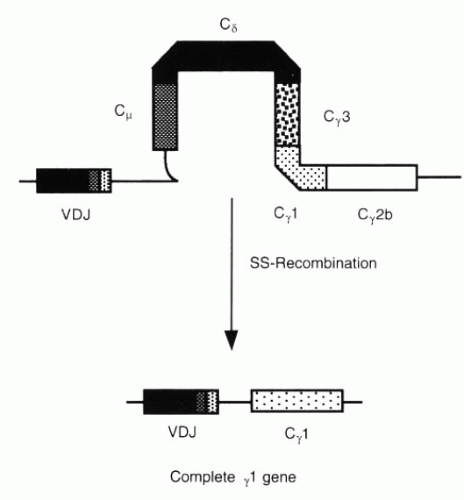 FIG. 3-10. The VHDJH gene moves from its position near Cμ to one near Cγ1 by SS recombination. (From Albert DA, Jakobiec FA. Principles and practice of ophthalmology, 2nd ed. Philadelphia: WB Saunders, 2000:69.) |
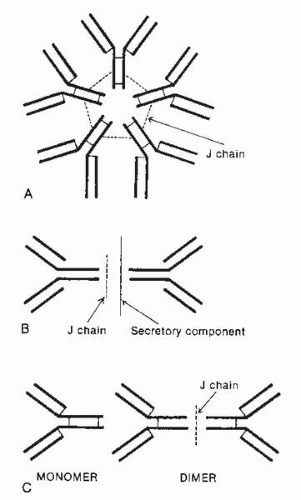 FIG. 3-11. Schematic diagram of polymeric human immunoglobulins. (From Albert DA, Jakobiec FA. Principles and practice of ophthalmology, 2nd ed. Philadelphia: WB Saunders, 2000:70.) |
and defend themselves against infection (46). The pathways of complement activation are complex (47) (Fig. 3-12). Activation begins with the formation of antigen-antibody complexes and the ensuing generation of peptides that leads to a cascade of proteolytic events. The particle that activates the system accumulates a protein complex on its surface that often leads to cellular destruction through disruption of membranes.
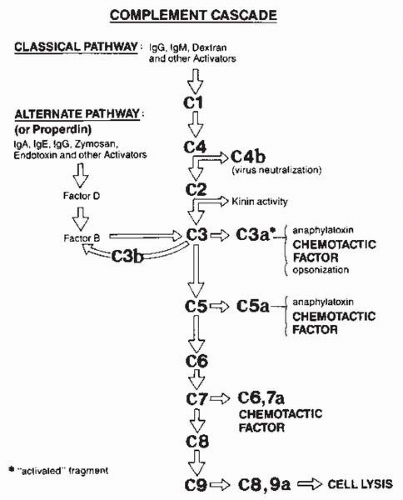 FIG. 3-12. Simplified schematic of steps in classic and alternate complement cascades. (From Albert DA, Jakobiec FA. Principles and practice of ophthalmology, 2nd ed. Philadelphia: WB Saunders, 2000:72.) |
maturation of the primary antibody response. Those cells with the greatest antigen-binding affinity survive as this primary B-cell response subsides, persisting as long-lived memory cells responsible for the classic distinguishing characteristics of secondary humoral immune response.
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


