Bacterial pathogens and microbiome alterations can contribute to the initiation and propagation of mucosal inflammation in chronic rhinosinusitis (CRS). In this article, the authors review the clinical and research implications of key pathogens, discuss the role of the microbiome, and connect bacteria to mechanisms of mucosal immunity relevant in CRS.
Key points
- •
The sinuses are not sterile. A population of bacteria is present in both health and disease, roughly in the same overall abundance, but qualitatively different in its makeup.
- •
As of yet, no single bacterial species or set of species has been definitively shown to be protective or causative in chronic rhinosinusitis (CRS).
- •
The overall function of the bacterial community may be most important, rather than the presence or absence of a single pathogen.
- •
Therapies used to treat CRS may induce microbiome alterations.
- •
Further research is indicated and required in this exciting field.
Introduction
Chronic rhinosinusitis (CRS) continues to be one of the most prevalent health care problems in the United States. Despite the significant morbidity, loss of productivity, and health care costs associated with CRS, the underlying processes that lead to disease remain poorly understood. The nonspecific clinical symptoms of nasal obstruction, rhinorrhea, facial pain, and anosmia may represent a common end point for various inflammatory mechanisms occurring in different anatomic areas. CRS is increasingly being appreciated as a clinical syndrome with a wide spectrum of overlapping disease physiology. For instance, CRS with nasal polyps (CRSwNP) often is characterized by eosinophilic inflammation and increased production of histamine, IL-5 and IL-13, whereas CRS without nasal polyps (CRSsNP) is often considered a predominantly neutrophilic disease characterized by high levels of interleukin (IL)-1, IL-6, and tumor necrosis factor (TNF) α. In practice, however, there are patients with CRSsNP with high levels of eosinophils and patients with CRSwNP who exhibit robust neutrophilic infiltration within the sinonasal epithelium. Thus, our classification of CRS in clinical practice is often not as simple as we would prefer.
Chronic Rhinosinusitis Pathophysiology and Immune Homeostasis
CRS is characterized by persistent inflammation, a dysregulated immune response, and host-microbial interactions that together result in disruption of epithelial barrier function, poor wound healing, tissue remodeling, and clinical symptoms. Historically the significance of bacteria in acute and CRS has focused on the interactions between a single bacterial pathogen and its host. However, developing concepts in microbial ecology, laboratory methods in culture-independent microbiology, and bioinformatics are furthering our capacity to study complex microbial communities as an entire functional unit. The nasal cavity and paranasal sinuses are the first tissues exposed to airborne environmental challenges, including pathogenic and nonpathogenic bacteria, viruses, fungi, allergens, and toxins. The mucosal surface uses several immune mechanisms to promote homeostasis, which can be broadly divided into innate and adaptive immunity. Many host factors impact the functionality of the immune response that is thought to predispose individuals to the development of CRS.
Innate versus adaptive immunity
Innate immunity classically refers to the nonspecific defense mechanisms that are rapidly activated following exposure to antigenic material and confer immediate protection. Within the upper respiratory tract, this includes the physical barrier provided by the ciliated pseudostratified columnar respiratory epithelium that lines the sinonasal cavity. This resilient barrier contains interspersed goblet cells that secrete a viscoelastic mucous layer atop the epithelial surface composed of high-molecular-weight glycoprotein mucins and heavily glycosylated molecules. In conjunction with beating cilia, the enriched mucous layer promotes nonspecific mucociliary clearance of microbes and irritant particles. Barrier dysfunction can contribute to CRS; when coupled with defects in mucociliary clearance that promote bacterial colonization, bacterial invasion and further barrier disruption may occur. Classic genetic defects in ciliary function, such as cystic fibrosis and primary ciliary dyskinesia, are often used as examples; but acquired ciliary dysfunction occurs in CRS as well. Poor barrier function and dysfunctional mucociliary clearance are host defects that predispose individuals to pathogen colonization and the development of recurrent infection.
Sinonasal epithelial cells secrete enzymes, opsonins, defensins, permeabolizing proteins, and other endogenous antimicrobial products into the apical mucous layer. These host defense molecules are important for directly neutralizing microbes and recruiting inflammatory cells that modulate the immune response. Specifically, epithelial cells secrete enzymes, such as lysozyme, peroxidases, and chitinases, and the small cationic permeabolizing proteins, such as the defensins and cathelicidins. Additionally, proteins, such as lactoferrin, mucins, C-reactive protein, and secretory leukocyte proteinase inhibitor, collectively provide protection from bacteria, fungi, and viruses at the apical surface.
When pathogens do invade the sinonasal epithelium, circulating professional phagocytes, possessing pattern recognition receptors (PRRs) on their cell surface, recognize pathogen-associated molecular patterns and damage-associated molecular patterns. When host epithelial cell PRRs bind pathogenic or damaged cell proteins, the acute inflammatory pathways are activated and the tissue becomes primed for the adaptive immune response. Toll-like receptors (TLRs) are a subset of PRRs that allow phagocytes to recognize bacterial motifs, such as lipopolysaccharide (via TLR-4) and CpG repeats (via TLR-9) associated with gram-negative bacteria and bacterial DNA respectively.
Current literature suggests that variable gene expression of TLRs and other cytokines may predispose some individuals to develop CRS. For example, patients with CRS demonstrate variable expression of TLR2 mRNA. TLR2 is expressed on sinonasal epithelial cells and recognizes several bacterial, fungal, and viral proteins. Binding of a ligand to TLR2 activates the acute inflammatory response and also primes the adaptive immune response by increasing expression of major histocompatibility complex II (MHC-II) costimulatory molecules required to present antigens to T-follicular helper cells. The resulting Th1 response is necessary for clearance of Streptococcus pneumoniae infection, for example. Another study identified decreased TLR9 mRNA expression in CRSsNP ; however, in-depth study of TLR-mediated disease in CRS is lacking.
In concert with the acute inflammatory response, cytotoxic natural killer (NK) cells are important to mounting an appropriate innate immune response. Although NK cells are often understood to induce apoptosis in viral-infected cells and tumor cells, they are becoming increasingly of interest in CRS. Patients with CRS have dysfunctional NK cells that demonstrate impaired degranulation and diminished release of TNF α and interferon (IFN) γ.
The adaptive immune system is stimulated when PMNs and macrophages are recruited to the site of infection to fully eradicate bacterial infection and establish long-lasting cell-mediated immunity. Antigen-presenting cells, such as monocytes, macrophages, B-lymphocytes, and dendritic cells, process complex protein antigens and present antigenic peptides on their surface that are then presented to T helper cells on the appropriate MHC. Binding of the antigen-MHC complex to the T-cell receptor activates the adaptive immune system and leads to longer-lasting T-effector cells and antibody-producing plasma cells.
Although much attention is given to defective innate immunity in the development of CRS, the adaptive immune system is crucial for developing an appropriate T-cell response; much of the CRS literature supports polarized populations of T-follicular helper cells (either Th1 or Th2) in subsets of CRS. Th1 cells produce IFN γ, IL-2, and TNF α, which ultimately lead to a robust cell-mediated response and phagocyte-mediated inflammation. Th2 cells produce IL-4, IL-5, and IL-13 that promote a strong antibody response and the accumulation of eosinophils while inhibiting the phagocytes and the resulting potent inflammatory response. Furthermore, the T-follicular helper cell environment influences macrophage differentiation into M1 macrophages that are necessary for mounting a vigorous inflammatory response against intracellular pathogens or M2 macrophages that are associated with eosinophilic states. M2 macrophages are predominant in CRSwNP and may play a role in exacerbating polyposis because they are unable to phagocytose Staphylococcus aureus . The inappropriate persistence of Th1 inflammation in CRSsNP and the ongoing recruitment of eosinophils and Th2 cells in CRSwNP are hallmarks of adaptive immune dysfunction characteristic in CRS.
Surface mucosal niche
Once thought to be a sterile environment in the healthy state, the paranasal sinuses are now widely appreciated to harbor rich and diverse populations of commensal bacteria. Commensals are most often defined as the community of microorganisms that colonize epithelial surfaces without causing harm to the host. Our understanding of commensal bacteria is evolving to recognize the shifting selective pressures in the microbial community that likely include other forms of symbiosis between host cells and neighboring bacteria including parasitism, mutualism and amensalism. Host defense mechanisms contribute to a unique mucosal environment that constantly applies selective pressure on epithelial microbes resulting in a surface mucosal niche. This niche varies between individuals and between different anatomic compartments. There may be drivers of this niche beyond host mucosal immune function, including environmental exposures (eg, smoking) or medication usage (eg, antibiotics) ( Fig. 1 ). Bacteria, either live or their by-products, such as DNA, quorum-sensing molecules, or metabolized waste, interact with the immune system and can influence inflammatory processes.
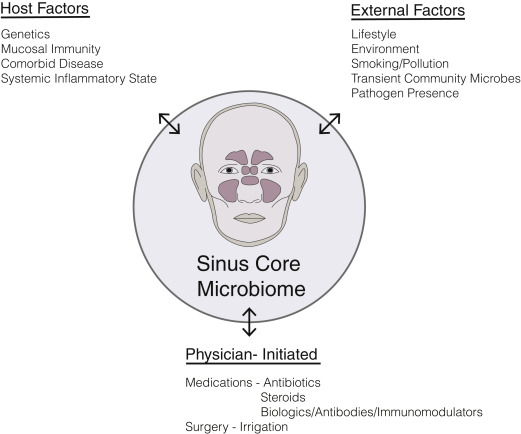
Role of Bacteria in Initiation and Sustenance of Inflammation
Historically, many described CRS as a disease that resulted from a persistent or incompletely treated acute infection. Current understanding is that CRS is an inflammatory process rather than infectious but that inflammatory mechanisms modulated by commensal and pathogenic bacteria contribute to disease formation, beyond the simplistic notion of a single microbial pathogen interacting with a host and causing disease. Certainly, chronic inflammatory diseases can result from direct bacterial invasion at mucosal surfaces, resulting in compromised barrier function, a coordinated innate and adaptive immune response, and an acute inflammatory response, which may evolve into prolonged inflammation leading to tissue damage, remodeling, and fibrosis. It is possible that in some patients, bacterial infection initiates the inflammatory process that never resolves; yet, in others it may be the case that a noninfectious inciting event initiates an inflammatory response that alters the native mucosal surface bacterial niche, which then propagates the disease once the original disease-causing event has concluded ( Fig. 2 ). Earlier studies of sinonasal microbiota used culture-based microbiology techniques, which have recently been usurped by nucleic acid–based molecular techniques that allow for more sensitive and less biased detection of microbes as well as the ability to characterize entire communities of microbes within the same sample. Without evidence to support a definitive role for bacterial infection as the cause for CRS, future studies are needed to better understand the role of bacteria in host susceptibility to disease development.

Introduction
Chronic rhinosinusitis (CRS) continues to be one of the most prevalent health care problems in the United States. Despite the significant morbidity, loss of productivity, and health care costs associated with CRS, the underlying processes that lead to disease remain poorly understood. The nonspecific clinical symptoms of nasal obstruction, rhinorrhea, facial pain, and anosmia may represent a common end point for various inflammatory mechanisms occurring in different anatomic areas. CRS is increasingly being appreciated as a clinical syndrome with a wide spectrum of overlapping disease physiology. For instance, CRS with nasal polyps (CRSwNP) often is characterized by eosinophilic inflammation and increased production of histamine, IL-5 and IL-13, whereas CRS without nasal polyps (CRSsNP) is often considered a predominantly neutrophilic disease characterized by high levels of interleukin (IL)-1, IL-6, and tumor necrosis factor (TNF) α. In practice, however, there are patients with CRSsNP with high levels of eosinophils and patients with CRSwNP who exhibit robust neutrophilic infiltration within the sinonasal epithelium. Thus, our classification of CRS in clinical practice is often not as simple as we would prefer.
Chronic Rhinosinusitis Pathophysiology and Immune Homeostasis
CRS is characterized by persistent inflammation, a dysregulated immune response, and host-microbial interactions that together result in disruption of epithelial barrier function, poor wound healing, tissue remodeling, and clinical symptoms. Historically the significance of bacteria in acute and CRS has focused on the interactions between a single bacterial pathogen and its host. However, developing concepts in microbial ecology, laboratory methods in culture-independent microbiology, and bioinformatics are furthering our capacity to study complex microbial communities as an entire functional unit. The nasal cavity and paranasal sinuses are the first tissues exposed to airborne environmental challenges, including pathogenic and nonpathogenic bacteria, viruses, fungi, allergens, and toxins. The mucosal surface uses several immune mechanisms to promote homeostasis, which can be broadly divided into innate and adaptive immunity. Many host factors impact the functionality of the immune response that is thought to predispose individuals to the development of CRS.
Innate versus adaptive immunity
Innate immunity classically refers to the nonspecific defense mechanisms that are rapidly activated following exposure to antigenic material and confer immediate protection. Within the upper respiratory tract, this includes the physical barrier provided by the ciliated pseudostratified columnar respiratory epithelium that lines the sinonasal cavity. This resilient barrier contains interspersed goblet cells that secrete a viscoelastic mucous layer atop the epithelial surface composed of high-molecular-weight glycoprotein mucins and heavily glycosylated molecules. In conjunction with beating cilia, the enriched mucous layer promotes nonspecific mucociliary clearance of microbes and irritant particles. Barrier dysfunction can contribute to CRS; when coupled with defects in mucociliary clearance that promote bacterial colonization, bacterial invasion and further barrier disruption may occur. Classic genetic defects in ciliary function, such as cystic fibrosis and primary ciliary dyskinesia, are often used as examples; but acquired ciliary dysfunction occurs in CRS as well. Poor barrier function and dysfunctional mucociliary clearance are host defects that predispose individuals to pathogen colonization and the development of recurrent infection.
Sinonasal epithelial cells secrete enzymes, opsonins, defensins, permeabolizing proteins, and other endogenous antimicrobial products into the apical mucous layer. These host defense molecules are important for directly neutralizing microbes and recruiting inflammatory cells that modulate the immune response. Specifically, epithelial cells secrete enzymes, such as lysozyme, peroxidases, and chitinases, and the small cationic permeabolizing proteins, such as the defensins and cathelicidins. Additionally, proteins, such as lactoferrin, mucins, C-reactive protein, and secretory leukocyte proteinase inhibitor, collectively provide protection from bacteria, fungi, and viruses at the apical surface.
When pathogens do invade the sinonasal epithelium, circulating professional phagocytes, possessing pattern recognition receptors (PRRs) on their cell surface, recognize pathogen-associated molecular patterns and damage-associated molecular patterns. When host epithelial cell PRRs bind pathogenic or damaged cell proteins, the acute inflammatory pathways are activated and the tissue becomes primed for the adaptive immune response. Toll-like receptors (TLRs) are a subset of PRRs that allow phagocytes to recognize bacterial motifs, such as lipopolysaccharide (via TLR-4) and CpG repeats (via TLR-9) associated with gram-negative bacteria and bacterial DNA respectively.
Current literature suggests that variable gene expression of TLRs and other cytokines may predispose some individuals to develop CRS. For example, patients with CRS demonstrate variable expression of TLR2 mRNA. TLR2 is expressed on sinonasal epithelial cells and recognizes several bacterial, fungal, and viral proteins. Binding of a ligand to TLR2 activates the acute inflammatory response and also primes the adaptive immune response by increasing expression of major histocompatibility complex II (MHC-II) costimulatory molecules required to present antigens to T-follicular helper cells. The resulting Th1 response is necessary for clearance of Streptococcus pneumoniae infection, for example. Another study identified decreased TLR9 mRNA expression in CRSsNP ; however, in-depth study of TLR-mediated disease in CRS is lacking.
In concert with the acute inflammatory response, cytotoxic natural killer (NK) cells are important to mounting an appropriate innate immune response. Although NK cells are often understood to induce apoptosis in viral-infected cells and tumor cells, they are becoming increasingly of interest in CRS. Patients with CRS have dysfunctional NK cells that demonstrate impaired degranulation and diminished release of TNF α and interferon (IFN) γ.
The adaptive immune system is stimulated when PMNs and macrophages are recruited to the site of infection to fully eradicate bacterial infection and establish long-lasting cell-mediated immunity. Antigen-presenting cells, such as monocytes, macrophages, B-lymphocytes, and dendritic cells, process complex protein antigens and present antigenic peptides on their surface that are then presented to T helper cells on the appropriate MHC. Binding of the antigen-MHC complex to the T-cell receptor activates the adaptive immune system and leads to longer-lasting T-effector cells and antibody-producing plasma cells.
Although much attention is given to defective innate immunity in the development of CRS, the adaptive immune system is crucial for developing an appropriate T-cell response; much of the CRS literature supports polarized populations of T-follicular helper cells (either Th1 or Th2) in subsets of CRS. Th1 cells produce IFN γ, IL-2, and TNF α, which ultimately lead to a robust cell-mediated response and phagocyte-mediated inflammation. Th2 cells produce IL-4, IL-5, and IL-13 that promote a strong antibody response and the accumulation of eosinophils while inhibiting the phagocytes and the resulting potent inflammatory response. Furthermore, the T-follicular helper cell environment influences macrophage differentiation into M1 macrophages that are necessary for mounting a vigorous inflammatory response against intracellular pathogens or M2 macrophages that are associated with eosinophilic states. M2 macrophages are predominant in CRSwNP and may play a role in exacerbating polyposis because they are unable to phagocytose Staphylococcus aureus . The inappropriate persistence of Th1 inflammation in CRSsNP and the ongoing recruitment of eosinophils and Th2 cells in CRSwNP are hallmarks of adaptive immune dysfunction characteristic in CRS.
Surface mucosal niche
Once thought to be a sterile environment in the healthy state, the paranasal sinuses are now widely appreciated to harbor rich and diverse populations of commensal bacteria. Commensals are most often defined as the community of microorganisms that colonize epithelial surfaces without causing harm to the host. Our understanding of commensal bacteria is evolving to recognize the shifting selective pressures in the microbial community that likely include other forms of symbiosis between host cells and neighboring bacteria including parasitism, mutualism and amensalism. Host defense mechanisms contribute to a unique mucosal environment that constantly applies selective pressure on epithelial microbes resulting in a surface mucosal niche. This niche varies between individuals and between different anatomic compartments. There may be drivers of this niche beyond host mucosal immune function, including environmental exposures (eg, smoking) or medication usage (eg, antibiotics) ( Fig. 1 ). Bacteria, either live or their by-products, such as DNA, quorum-sensing molecules, or metabolized waste, interact with the immune system and can influence inflammatory processes.
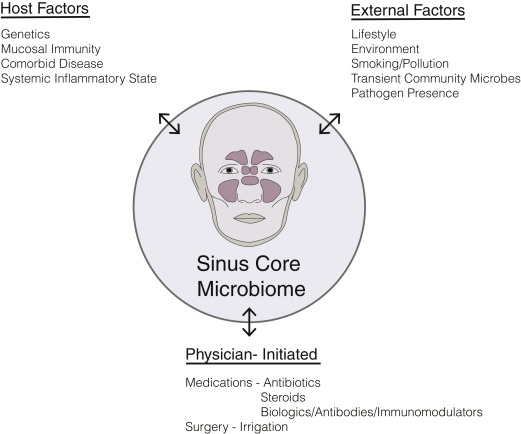
Role of Bacteria in Initiation and Sustenance of Inflammation
Historically, many described CRS as a disease that resulted from a persistent or incompletely treated acute infection. Current understanding is that CRS is an inflammatory process rather than infectious but that inflammatory mechanisms modulated by commensal and pathogenic bacteria contribute to disease formation, beyond the simplistic notion of a single microbial pathogen interacting with a host and causing disease. Certainly, chronic inflammatory diseases can result from direct bacterial invasion at mucosal surfaces, resulting in compromised barrier function, a coordinated innate and adaptive immune response, and an acute inflammatory response, which may evolve into prolonged inflammation leading to tissue damage, remodeling, and fibrosis. It is possible that in some patients, bacterial infection initiates the inflammatory process that never resolves; yet, in others it may be the case that a noninfectious inciting event initiates an inflammatory response that alters the native mucosal surface bacterial niche, which then propagates the disease once the original disease-causing event has concluded ( Fig. 2 ). Earlier studies of sinonasal microbiota used culture-based microbiology techniques, which have recently been usurped by nucleic acid–based molecular techniques that allow for more sensitive and less biased detection of microbes as well as the ability to characterize entire communities of microbes within the same sample. Without evidence to support a definitive role for bacterial infection as the cause for CRS, future studies are needed to better understand the role of bacteria in host susceptibility to disease development.
The paranasal sinus biome in health and disease
The Microbiome
Host-microbe interactions are an established contributing factor in the formation of CRS, but evidence for the presence or absence of a single microbe resulting in disease is lacking. The shifting paradigm focuses on the unique composition of the entire bacterial community that colonizes the sinonasal mucosa also known as the microbiome. The microbiome is a potentially diverse community of microbiota existing in a delicate symbiotic relationship within a human microenvironment. These organisms possess great genetic potential to act as disease modifiers and contribute to the maintenance of health and formation of disease on all epithelial surfaces, including upper and lower airway. In intestinal epithelia, early microbial colonization is essential for normal immunologic development and influences susceptibility to inflammatory and allergic diseases later in life. In neonates, for example, early lower airway colonization with pathogens, such as S pneumonia, Haemophilus influenzae , and Moraxella catarrhalis , increases the risk of recurrent wheezing and asthma. Additionally, several groups have demonstrated the importance of the microbiome on the host adaptive immune system through modulation of dendritic cells, Th17, and T-regulatory (Treg) cells. Although much of bacterial microbiome research is still emerging, it is one of the most intriguing current topics of CRS research.
There remains a paucity of microbiome research on fungi, viruses, and bacteriophages in CRS, although growing evidence suggests that these microbes may contribute to a larger meta-organism in which interactions between and among all microbes and their host shape human health. Early efforts to describe microbial roles in CRS resulted in the fungal hypothesis, which suggested that CRS resulted from an overexuberant host response to Alternaria fungi. Although this theory does not explain many of the defects observed in CRS, and current guidelines recommend against antifungal therapy for patients with CRS, there is continued research interest in the fungal microbiome. Recent studies using sequencing of the fungal 18S ribosomal RNA gene demonstrate a rich and diverse population of commensal fungal taxa in middle meatal lavages from healthy patients and patients with CRS. Patients with CRS were found to have quantitative increases in the total amount of fungal 18S ribosomal RNA when compared with controls. A 2014 prospective study by Cleland and colleagues identified 207 unique fungal genera in 23 patients with CRS and 11 controls. Interestingly, fungal genera traditionally associated with CRS, such as Alternaria and Aspergillus , were found in very low abundance. This study also assessed postoperative changes in the fungal microbiome and found decreased richness at 6 and 12 weeks after surgery. No studies to date have thoroughly profiled viral or bacteriophage populations within the paranasal sinuses, although there is certainly interest. For instance, recent evidence suggests that upper airway rhinovirus infection can alter the nasopharyngeal microbiome. Further studies directed at characterizing the entire microbiome population in health are necessary in order to develop a more robust understanding of the role microbial diversity plays in sinonasal health as well as the generation of disease.
Bacteria in Health
Since the advent of culture-independent molecular techniques, several groups have used various techniques to characterize the sinonasal bacterial community in healthy individuals. Efforts to identify a distinct microbial profile in health are inconclusive, and there is currently no consensus on which bacteria predominate in or define the healthy state. Although several studies have characterized the bacterial communities in the upper airway, cross-study comparisons are difficult because of several variations in sampling methods, bacterial primer selection, sequencing methods, and data analysis pipelines. Even so, there are several patterns that frequently emerge. These patterns include an abundance of Propionibacterium acnes, Staphylococcus epidermidis, S aureus, and Corynebacterium spp in health. Of note, Staphylococcus spp, including S aureus and coagulase negative staphylococci, are present in healthy subjects and can behave in either pathogenic or commensal fashion based on particular strain, bacterial gene expression, and surrounding microbial interactions. Although native bacteria act to promote homeostasis within the sinonasal epithelia, disruption of this balance, known as dysbiosis , may allow commensal organisms to act as opportunistic pathogens in disease states.
The Human Microbiome Project (HMP) consortium compiled 16S rRNA gene sequences collected from 18 different sites across the body in order to better associate the microbiome with human health. Further analysis of the HMP data demonstrates that different human subjects possess unique native bacterial communities with highly variable taxonomic composition at the same body sites. These bacterial community profiles strongly associated with life-history characteristics, such as history of being breastfed as an infant, sex, and level of education. Interestingly, despite profound interperson and intraperson variability in the bacterial microbiome, community function remains constant; the community types from one body site are predictive of community types at other body sites within the same individual. The association of bacterial community composition with life history factors, such as presence or absence of breastfeeding, raises questions about the source of commensal microbes. In the gastrointestinal literature, there are several studies that demonstrate that diet shapes the microbiome composition and different microbial profiles are associated with diseases, such as inflammatory bowel disease (IBD). In addition to dietary factors, the upper airway microbiota are a known source of microbes that colonize the lung and stomach. Ultimately, the aggregate of bacterial taxa in the sinus niche need to be better studied at a metagenome level to begin deciphering community function and how specific bacterial ecosystems are established.
Microbiome Changes in Disease
Several statistical indices are use to describe species diversity within a microbial community. Alpha diversity is the intracommunity diversity as measured by the total number and composition of species. Beta diversity is an estimate of the diversity (number and composition) comparison between different habitats. Abundance refers to the total amount of bacterial DNA that corresponds to specific bacterial taxa found within a sample. Richness describes the total number of species identified within a sample, that is, greater numbers of distinct species means higher richness. Evenness is a measure of how similar in number each species within a bacterial community is. Several studies have identified perturbations in the microbiome during CRS. In general, S aureus and anaerobes, including Prevotella, Fusobacterium, Bacteroides spp, and Peptostreptococcus spp, are consistently more abundant in CRS versus healthy controls. Interestingly, although the abundance of pathogenic bacteria is increased in patients with CRS, the overall total quantity of bacteria does not seem to change compared with healthy individuals. This finding indicates that the sinus niche is filled by a given amount of bacteria, and a relative dominance of the niche by pathogens is associated with disease. In these studies, despite having a similar overall bacterial burden, microbial richness, evenness, and diversity of bacterial colonies were greatly diminished in CRS. This observation supports the hypothesis that disturbances in the microbial community are a part of CRS. Just as ecologists associate the macroscopic biodiversity and biomass of rainforests and coral reef habitats with the overall health of the ecosystem, the microscopic biodiversity of bacteria within a host mucosal niche is considered a hallmark of health.
The frequent observation that CRS is associated with decreased microbial diversity parallels many other disease states. For example, decreased diversity in the gut microbiome is associated with obesity, active IBD, and psoriatic arthritis. Although data are lacking to suggest that promoting bacterial diversity within the sinonasal niche would prevent or improve CRS, this may be a relevant consideration moving forward because the mainstays of CRS therapy are long-term broad-spectrum antibiotics and corticosteroids, which carry the potential to alter the microbiome composition.
Are Microbiome Alterations a Cause or By-product of Disease?
Recent cross-sectional population studies have detected differences in the microbiome between patients with CRS and healthy individuals, but what does this really mean? One possibility is that community alterations in the microbiome lead to epithelial barrier and immune dysfunction resulting in disease. Alternatively, persistent inflammation at the mucosal surface may result in a prolonged immune response and/or alterations in the local microenvironment that shift the microbial community. Moreover, therapies administered for the disease (eg, antibiotic therapy and steroids) likely impact the microbiome and may confound these observations. The multitude of variables present that impact the mucosal niche makes establishing this causation difficult in the absence of animal models. In reality, any or all of these possibilities may occur (see Fig. 1 ).
The concept of perturbations in the microbial community resulting in disease, that is, dysbiosis , has been explored in the gastrointestinal tract. The normal gut flora plays an essential role in maintaining immune homeostasis, and disruptions in these mutualistic relationships are thought to lead to several conditions ranging from antibiotic-associated diarrhea and IBD to necrotizing enterocolitis and colorectal cancer. Although disease-associated shifts in the bacterial microbiome have been observed in CRS, it remains unclear if this is an inciting factor in the development or propagation of disease or merely a consequence. Emerging data from cross-sectional analysis of specific patient populations suggest that host factors, such as a history of tobacco use, presence of asthma, or recent antibiotic use, can impact the microbiota.
Given the panoply of disease mechanisms involved, the prolonged utilization of medical therapies, and the lack of a universal mouse model for the disease, human studies of CRS will require large-scale, multilevel, longitudinal design in order to delineate patterns of microbial perturbations and their significance. Such studies are critical to determining the role of the microbiome in disease formation, chronicity, severity and prognosis, and response to therapies.
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


