Fig. 3.1
Formation of the pharyngeal pouches and their corresponding derivatives. From [2]. PA pharyngeal arch
These endodermal outpockets contact the surface ectoderm at precise sites along the anterior–posterior axis of the pharyngeal apparatus, and they expand along the dorsoventral axis to generate their characteristic narrow, slit-like shape [4, 5]. The two most cranial pouches , 1 and 2, form first followed by pouches 3 and 4 [4, 6]. The parathyroids glands develop from the most caudal pouches , but the number of glands varies depending on species [3]. In humans and birds, two sets of parathyroids (from the third and fourth pouches) develop, while only one set (from the third pouch) forms in rodents [3]. In humans, the epithelium of the dorsal surface of the third pouch differentiates into the inferior parathyroid glands, while the ventral surface differentiates into the thymus [2]. The epithelium of the dorsal wing of the fourth pouch differentiates into the superior parathyroid glands, while the ventral wing differentiates into the ultimobranchial body (Fig. 3.1; [7, 8]).
Using murine and chick-quail models , much progress has been made in identifying specific transcription factors (Hoxa3-Eya1-Pax1/9-Six1/4 and Tbx1) and signaling molecules (Bmp4, Fgf8, Shh, Wnt5b) involved in the early formation and patterning of the third and fourth pouches [9–11]. The pouches are highly polarized structures delineated into precise molecular domains [4, 6, 12, 13]. In the third pouch, the epithelial cells express either Gcm2, destined to become parathyroid tissue, or Bmp4 and Foxn1, destined to become thymus [10, 12–14]. In the chick, Gcm2 is expressed in both the third and fourth pouches, supporting parathyroid development from both of these pouches [11]. Of note, Gcm2 is not required for early third pouch patterning into thymus/parathyroid domains, but appears to be required for later parathyroid differentiation and survival [13]. Thus, the organ-specific cell types are already established even before expression of Gcm2, as indicated in the thymus by immigration of lymphocyte progenitors and in the parathyroid by expression of PTH, P75NTR, S100 protein, and chromogranin A [7, 13, 15]. The primary induction cue that defines whether the third pouch precursor cells are destined to become parathyroid or thymus remains unknown [10, 13].
In humans, a number of hypoparathyroid developmental anomalies have been investigated, revealing important roles for particular genes involved in early third and fourth pouch morphogenesis. In DiGeorge syndrome (DGS) , there is congenital failure in the development of the third and fourth pouch derivatives with resulting hypoplasia or agenesis of the parathyroid glands and thymus [8]. These patients may present with a range of findings: hypoparathyroidism with hypocalcemia, immune deficits secondary to lack of T-cell function, cardiac defects including truncus arteriosus, and characteristic facies [16]. Microdeletions mapped to chromosome 22q11.2 (referred to as DGSI) have been described in up to 90 % of these patients [8, 16]. At present, the gene thought to be responsible for DGSI is TBX1. TBX1 is a T-box transcription factor that is expressed in highly regionalized domains during early pharyngeal development. In murine models, Tbx1 mutants have markedly decreased expression of Gcm2, suggesting that this parathyroid-specific marker is one of the downstream targets of the DGS1 gene [8].
Similarly, abnormal expression of GCM2 has been identified in other parathyroid-related disorders. Homozygous inactivating mutations in GCM2 have been described in human familial autosomal recessive and dominant forms of isolated hypoparathyroidism. These individuals may have undetectable PTH serum levels. It is not yet clear whether the parathyroids are unformed or hypoplastic in this population [17, 18]. In addition, there have also been reports of parathyroid adenomas associated with deregulated (both elevated and reduced) expression of GCM2. At present, the clinical significance of these findings remains unclear [19, 20]. In any case, this transcription factor required in both differentiation and ultimate survival of the parathyroid glands is providing new insights into how the parathyroid gland forms and functions.
Separation from the Pharynx
The inferior and superior parathyroid glands detach from the pharyngeal wall around the end of the fifth gestational week [21]. The mechanism by which the thymus and parathyroid primordia detach from the pharynx has been extensively studied using murine models [22]. Early in development, the site of pharyngeal attachment is shared equally between the Gcm2+ parathyroid and Bmp4+ thymus domains [7, 22]. Neural crest cells , surrounding the pouches, express molecular signals that define the border location and size of the two molecular gradients [7]. Preferential proliferation of the thymus primordium relative to the parathyroid primordium, however, results in a dramatic increase in size and caudal growth of the thymus [23]. This unequal growth pattern results in the parathyroid domain, located on the cranial portion of the thymus, being displaced from its pharyngeal attachment [24]. Thus, the thymus persists as the sole connection to the pharynx [7, 22]. Apoptotic epithelial cells have been identified at the final attachment site, suggesting that separation is mediated by programmed cell death. The molecular signaling pathway directing this site-specific apoptosis remains elusive. However, recent evidence suggests that low levels of FGF signaling, which is associated with cell-mediated death in several other developmental contexts, may be involved [22].
Separation from the Thymus
A number of mutations in both the neural crest and endoderm have been associated with delayed thymus and parathyroid separation, suggesting that the interaction between these two cell types is key [14, 23–25]. During embryogenesis , the pharyngeal organs, especially the thymus, are densely surrounded by mesenchyme derived from the neural crest [26]. Recent evidence suggests that the neural crest mesenchymal cells migrate in between the Bmp4+ and Gcm2+ epithelial domains and act as a physical “wedge” separating the two zones [14].
Based on observations in both mice and humans, separation is frequently incomplete [27, 28]. In murine models, using high-resolution in situ hybridization , the Gcm2+ parathyroid domain shows a relatively high tendency to fragment and leave small parathyroid clusters either attached to or trailing the thymus into the mediastinum [7, 27]. It is thought that the generation of these microscopic parathyroid clusters is the source of supernumerary or accessory parathyroid tissue [7, 27]. In humans, the prevalence of supernumerary glands is between 2 and 6 % [29], and as many as 11 glands have been reported in large autopsy series [28, 30]. In 2/3 of cases, the fifth gland is inferior to the lower pole of the thyroid associated with the thyrothymic ligament or the thymus. The remaining third of supernumerary glands are typically adjacent to the thyroid between the orthotopic superior and inferior parathyroids [30].
A number of studies have suggested that the thymus, which shares such an intimate origin with the parathyroid gland, may serve as an auxiliary source of PTH [27]. In murine models , extra-parathyroid PTH production has been reported from misplaced, isolated parathyroid cells that were most commonly still attached to the thymus [27]. In addition, medullary thymic epithelial cells express PTH in a Gcm2-independent manner as a means for self-selection [27]. However, at present, it does not appear that these medullary cells provide any auxiliary endocrine function and the clinical significance remains elusive [27]. Instead, the generation of multiple, microscopic parathyroid clusters is the most likely source of any physiologically relevant “thymic PTH.”
Migration
In humans, the inferior and superior parathyroid glands migrate inferiorly and medially until arresting on the dorsal side of the caudal thyroid lobes around the seventh week [31]. Migration of the pharyngeal derived organs has been particularly well characterized in murine models. Following detachment, the thymus/parathyroid primordium moves in a caudal-ventral-medial direction towards the anterior thoracic cavity [2]. During this time, the parathyroid domain remains attached to the cranial pole of the thymus [2]. When the thymus passes along the lateral sides of the thyroid, the parathyroids detach and remain adjacent to the thyroid [2].
The thymus, in contrast, continues its migration into the anterior mediastinum where it joins its contralateral thymic lobe [2]. Following separation of the parathyroids from the thymus, the parathyroids do not migrate any further caudally [7]. At present, there is no evidence that the parathyroids are able to migrate independently of the thymus lobes [7]. Interestingly, the parathyroids lack a well-defined mesenchymal capsule, which is thought to be the source of the migrating neural crest driving thymus migration [7]. The variable location of the inferior glands in the neck then is likely a consequence of wherever they separate from the thymus [7]. In humans, when no migration occurs, the inferior glands remain at the level of the third pharyngeal pouch, near the bifurcation of the carotid artery where it appears to be located higher (and more ventral) than its superior counterparts [21].
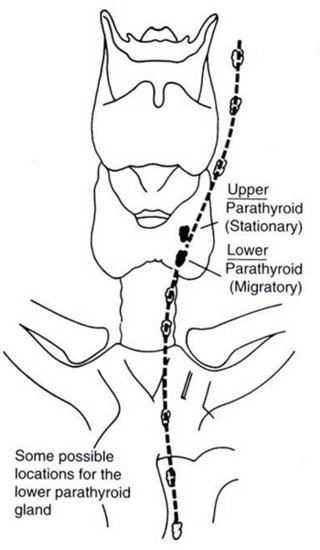
The long descent of the inferior parathyroid glands with the thymus from the neck into the anterior mediastinum is responsible for their highly variable location, which may be anywhere from the hyoid bone to the lower mediastinum (Fig. 3.2). In 50 % of cases, the inferior parathyroid gland may be found within 1 cm inferior, lateral or posterior to the inferior pole of the thyroid [28]. It is typically anterior to the coronal plane drawn along the recurrent laryngeal nerve. The recurrent laryngeal nerve courses around the fourth aortic arch in between the ventral side of the superior glands and the dorsal aspect of the inferior glands [32].

Fig. 3.2
Migratory descent of the inferior parathyroid glands during embryogenesis may result in highly variable anatomic location, which may be anywhere from the angle of the mandible to the pericardium. From http://www.thyroidmanager.org/wp-content/uploads/2011/06/35-8.png
In a large autopsy series (N = 942) including 3796 parathyroid glands, 324 glands (8.5 %) were ectopic. The most common sites for surgical exploration of ectopic inferior glands are anywhere along the common carotid artery, from the level of thyroid to the pericardium [29]. The most common ectopic location for an inferior gland is in the anterior mediastinum , representing some 5–6 % of ectopic cases [28, 29]. The glands are found in the upper mediastinum almost twice as frequently than in the lower mediastinum [29]. In young adults, the glands may be included in the lobular remnants of the thymic horns [21]. During embryogenesis , the inferior glands may also be displaced posterior and inferiorly resulting in their location along the recurrent laryngeal nerve on the lateral border of the esophagus. Further, rarely, they may be displaced posterior and laterally resulting in their unusual location in the upper posterior mediastinum [21].
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree



{kind=link}