Anatomy and Physiology of the Retina and Optic Nerve: Distinguishing Retinal from Optic Nerve Disease
Knowing the anatomy and physiology of the retina and optic nerve is crucial in understanding the similarities and differences in findings in patients with retinal versus optic nerve disease.
Anatomy and Physiology of the Retina
The retina is a unique part of the nervous system because it can be visualized, allowing the clinician to observe directly the effects of a wide range of diseases such as an infarction in evolution, deposition of metabolic storage products, or slowing of axoplasmic flow. The retina is also a favorite tissue for study by neuroscientists because it is thin and can be dissected easily from the eye because its cells are segregated into layers, and because there is at least a fundamental appreciation of its anatomic and physiologic organization.
The cellular structure of the retina is enormously complex. In humans, it contains over 100 million neurons, among which are about 30 different cell types that use at least 10 different neurotransmitters. Most architectural schemas underrepresent the intricacy of the retina because the degree to which information is shared among various cells depends on a variety of factors, including the region of the retina being illuminated, the characteristics of the light stimulus, and the state of adaptation of the retina to light. Given this complexity, one might imagine that a large number of symptoms would result from dysfunction of these many cell types. On the contrary, most clinically recognized diseases of the retina are caused by dysfunction of either photoreceptors or retinal ganglion cells (RGCs).
The human retina covers an area approximately 2500 mm2 and extends from the ora serrata to the optic disc. The retina contains six classes of neurons (photoreceptors, horizontal cells, bipolar cells, amacrine cells, interplexiform cells, and RGCs) and two types of glial cells (astrocytes and Müller cells) that are arranged into three parallel layers, except in the perifoveal zone, where the retina thins to a single layer (Table 2.1; Figs. 2.1 and 2.2). The human retina is roughly 120 μm thick over most of its area, with a maximum thickness of 230 μm in the macula and a minimum thickness of 100 μm in the foveal pit.
Cell Types and Layers of the Retina
The retinal pigment epithelium (RPE) is a monolayer of cells derived from the outer wall of the optic vesicle. It is intimately associated with the outer segments of the photoreceptors (Fig. 2.1). The RPE is responsible for several essential functions. Most significantly, it (a) is integrally involved in the phototransduction cascade through regeneration of chromophore; (b) provides trophic support for photoreceptors; (c) is a transport barrier between the choriocapillaris and the sensory retina; (d) participates in ocular immune responses; (e) binds reactive oxygen species; and (f) absorbs excess radiation with their melanin granules.
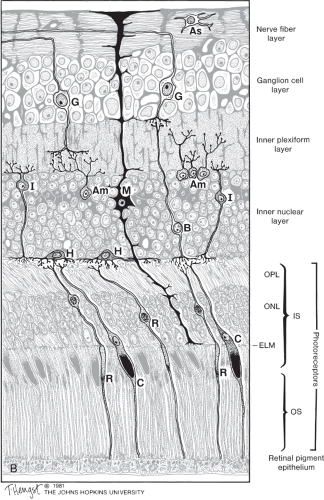 Figure 2.1 Cellular components of the fully developed primate retina, showing cellular relationships and major synaptic interactions. OPL, outer plexiform layer; ONL, outer nuclear layer; ELM, external limiting membrane; OS, outer segment; As, astrocyte; G, ganglion cell; Am, amacrine cell; I, interplexiform cell; H, horizontal cell; B, bipolar cell; C, cone; R, rod; M, Müller cell. The photoreceptor nuclei lie in the ONL, and a large stretch of the photoreceptors consists of their outer segments, which lie proximal (i.e., nearer to the retinal pigment epithelium) to their nuclei. The outer segments contain chromophore, which captures the energy from incoming light and initiates the ionic exchange that hyperpolarizes the photoreceptors. The inner segments of photoreceptors (not labeled) are the segments between the outer segments and the nucleus that contain a high concentration of mitochondria and endoplasmic reticulum. |
Table 2.1 Cellular Organization of the Retina | ||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
|
The visual cycle begins when the outer segment of a photoreceptor absorbs a photon, causing isomerization of 11-cis-retinol, the dark adapted form of rhodopsin, to all-trans-retinol. This reaction initiates the phototransduction cascade that ultimately influences propagation of nerve impulses through the retina (Fig. 2.3). The 11-cis-retinol must be continuously regenerated to maintain the ability of the photoreceptors to respond to light, and this regeneration is performed by the RPE. Patients in whom regeneration of photopigment is abnormally slow have a prolonged recovery of central vision following exposure of the retina to bright light. This phenomenon is present in patients with various macular dystrophies, age-related macular degeneration, and central serous chorioretinopathy, but also in patients with severe stenosis of the ipsilateral internal carotid artery causing markedly reduced oxygen delivery to the photoreceptors. It can be detected clinically by performing a photostress test (see Chapter 1).
The second primary function of the RPE—membrane renewal—is just as important as RPE’s role in phototransduction. Outer segments of rods and cones are shed on a daily basis. The outer segment of a human rod is entirely replaced over a period of 10 days and, although the timing of cone shedding is more variable, the involvement of the RPE in this process of shedding and renewal is integral. Disruption of the interaction between the RPE and the photoreceptors can lead to retinal degeneration, as occurs in hydroxychloroquine toxicity.
The photoreceptor cells of the retina are the sensory transducers for the visual system. They convert electromagnetic energy (i.e., light) into a neural signal. The photoreceptors contain visual pigments that, by absorbing photons of light, initiate the phototransduction process. These pigments absorb within the range of electromagnetic wavelengths that pass through the cornea and lens, generally between 400 and 700 nm. The two types of photoreceptors are the rods and the cones.
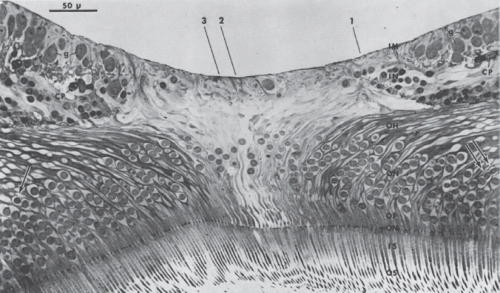 Figure 2.2 Section through center of fovea. Nuclei of rod receptor cells are indicated by arrows. Remaining receptor cells are foveal cone cells. OS, outer nuclear layer; OF, outer cone fiber; OH, outer fiber layer of Henle; IN, inner nuclear layer; g, ganglion cell; CP, capillary; IM, internal limiting membrane; IS, inner segment; OM, outer limiting membrane; ON, outer nuclear layer. (Reduced from ×400.) (Reprinted with permission from Yamada E. Some structural features of the fovea centralis in the human retina. Arch Ophthalmol. 1969;82:151–159.) |
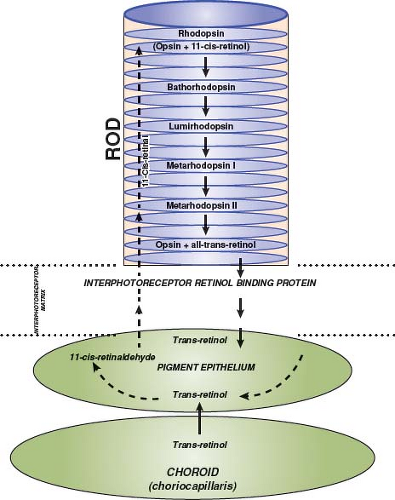 Figure 2.3 Phototransduction cascade. Schematic drawing of junction of rod, retinal pigment epithelium, and choroid to show the interrelationship among those structures required to regenerate photopigment. trans-Retinol is carried by the blood to the choriocapillaris where it can then enter the retinal pigment epithelium and contribute to the cycle. |
Both rods and cones have inner and outer segments, both of which are located outside the boundary of the outer limiting membrane (Fig. 2.4). The inner segment contains the ellipsoid, a structure that is richly endowed with mitochondria. The ellipsoid is responsible for the high level of oxidative metabolism that occurs in photoreceptors. The myoid is the region between the ellipsoid and the nucleus of the photoreceptor. This region contains numerous cellular organelles. The outer segment is connected to the inner segment by a cilium, which contains nine pairs of microtubules arranged in a pattern that is characteristic of nonmotile cilia.
Rods contain rhodopsin, a molecule that is composed of the apoprotein, opsin, and 11-cis-retinol. Rhodopsin is present in very high concentrations within the membranes of the outer segment discs, which are oriented vertically to maximize capture of incident photons. Activation of the phototransduction cascade causes an intracellular decline in cyclic guanidine monophosphate (GMP) concentration within the rod outer segment, followed by closure of the cyclic GMP-gated cation channel and cessation of inflow of extracellular cations that normally generates the dark current. The rod becomes hyperpolarized upon exposure to light, which decreases the amount of neurotransmitter released at the synaptic terminal.
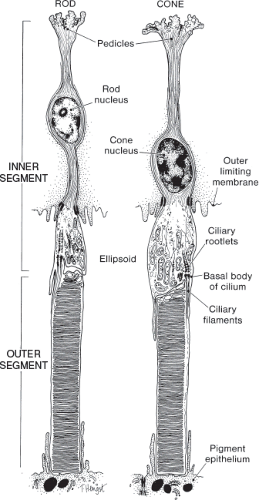 Figure 2.4 Ultrastructure of photoreceptors. Schematic drawing of photoreceptor cells showing the relationship of the inner and outer segments. The ellipsoid is located at the apex of the inner segment. (Reprinted with permission from Hogan MJ, Alvarado JA, Weddell JC. Histology of the Human Eye: An Atlas and Textbook. Philadelphia, PA: WB Saunders; 1971.) |
Vitamin A is used in the process of phototransduction and regeneration of photopigment. Vitamin A must be replenished from the circulation, and failure to do so can result in a bilateral retinopathy that clinically may mimic a bilateral optic neuropathy; however, because rods are predominately affected in the retinopathy caused by deficiency of vitamin A, nyctalopia (night blindness) usually is the initial manifestation of this disorder. This symptom helps to distinguish
the retinopathy from an optic neuropathy. Hypovitaminosis A is a highly significant problem in developing countries, but also occurs in persons who live in developed countries but who have malabsorption syndromes or liver disease, who adhere to restrictive diets, or who have poor diets due to chronic alcohol abuse.
the retinopathy from an optic neuropathy. Hypovitaminosis A is a highly significant problem in developing countries, but also occurs in persons who live in developed countries but who have malabsorption syndromes or liver disease, who adhere to restrictive diets, or who have poor diets due to chronic alcohol abuse.
Cones also contain 11-cis-retinol but have different apoproteins than rods. The variation of the opsin moieties produces three different spectral response curves corresponding to the blue, green, and red cones, which are more appropriately called short- (S), medium- (M), and long- (L) wavelength cones to emphasize that their responsiveness is not limited to a single color but rather is spread across a portion of the visible spectrum. Congenital color blindness almost always results from defects in either the red or green photopigment gene, both of which are located on the X chromosome.
Humans have a cone:rod ratio of approximately 1:20, with each retina containing 140 million rods and 7 million cones. About 50% of the cones are located within the central 30 degrees of the visual field, an area that roughly corresponds to the size of the macula. Rods are absent in the fovea and are in highest density in an elliptical ring, the center of which is the optic nerve head. There is an age-related decline in the number of rods in the human retina, with an annual loss of about 0.2% to 0.4%.
The density of foveal cones poses a problem in regard to their forward connections because their synaptic terminals (called cone pedicles) and the ganglion cells to which they connect have larger diameters than the cone inner segments themselves. Thus, each foveal ganglion cell soma is displaced away from the site of the cone that initiates its visual response. This realignment is accomplished by the nerve fiber layer (NFL) of Henle, an elongation of cones and rods between their nuclei and synaptic terminals.
The “midget” system refers to the anatomic pattern of cone outflow within the most central retina, although there is no specific boundary beyond which the midget pathway cannot be found. The midget system provides the most direct communication of spatial detail from outer to inner retina. In this system, output from one cone passes to one ON-center and one OFF-center bipolar cell with each bipolar cell connecting in turn to a single ON- or OFF-center ganglion cell. Thus, the divergence of signal from a foveal cone to a ganglion cell is 1:2. In the peripheral retina, by comparison, there is marked convergence of information from outer to inner retina, with perhaps up to 1500 rods influencing the function of one ganglion cell.
The outer plexiform layer (OPL) is the zone of synaptic connection between cells whose nuclei are in the outer and inner nuclear layers (Fig. 2.1). Cone pedicles and rod spherules, the synaptic specializations of cones and rods, respectively, have several invaginations into which processes of horizontal and bipolar cells extend, creating “triads” of one bipolar cell and two horizontal cells.
The retina contains two types of horizontal cells (Fig. 2.1). The HII cell, the smaller of the two types, makes dendritic contact with cones and axon contact with rods, whereas both ends of the HII cell synapse only with cones. Horizontal cells seem to provide the anatomic substrate for the center-surround response characteristics observed in the physiologic recordings made from ganglion cells and some other inner retinal neurons. Specifically, the response of a ganglion cell to light that falls within the center of its receptive field is opposite to that, which occurs to light that falls in the surround (i.e., the portion of the receptive field beyond the center). The antagonistic center-surround organization emphasizes contrast within the visual scene, which enhances spatial resolution.
Bipolar cells receive input from photoreceptors and provide output to both amacrine cells and ganglion cells (Fig. 2.1). Some bipolar cells receive input from rods, others from cones. Rod bipolar cells are the target of an apparent paraneoplastic immune attack in some patients with cutaneous malignant melanoma, so-called melanoma-associated retinopathy (MAR). The physiologic consequence of destruction of these cells is loss of the scotopic b-wave of the electroretinogram (ERG).
The major excitatory neurotransmitter in the retina is glutamate, and this substance has a profound effect at the level of the bipolar cells. There are two physiologic types of bipolar cells: hyperpolarizing (OFF-center cells) and depolarizing (ON-center cells). These bipolar cells respond differently to changes in the ambient glutamate concentration within the OPL. Release of glutamate by photoreceptors initiates two opposing but parallel signals to the inner retina (Fig. 2.5).
Amacrine cells are so named because they lack axons. Their cell bodies are located primarily in the inner nuclear layer of the retina (Fig. 2.1). Some amacrine cells are interneurons between bipolar and ganglion cells. Others connect to fellow amacrine cells, forming a lateral pathway within the inner plexiform layer (IPL) similar to that formed by horizontal cells in the OPL. There are numerous subtypes of amacrine cells.
The cell bodies of the interplexiform cells reside in the IPL (Fig. 2.1). These cells are postsynaptic to amacrine and bipolar cells in the IPL and presynaptic to horizontal and bipolar cells. This cell thus appears to be unique in that it apparently conveys information from the inner to the OPL, against the standard direction of neural transmission within the retina.
The retina contains several types of glial cells that have various functions. The most important are Müller
cells and astrocytes. Müller cell somas are located in the inner nuclear layer, but processes from these cells extend throughout the entire retina (Fig. 2.1). The apical processes of Müller cells form the outer limiting membrane by making junctional complexes with one another and with photoreceptors. On the vitread side, apposed Müller cell end feet forms the inner limiting membrane. Lateral extensions from Müller cells contact neurons within cellular layers, synapses within plexiform layers, axons within the NFL, and blood vessels.
cells and astrocytes. Müller cell somas are located in the inner nuclear layer, but processes from these cells extend throughout the entire retina (Fig. 2.1). The apical processes of Müller cells form the outer limiting membrane by making junctional complexes with one another and with photoreceptors. On the vitread side, apposed Müller cell end feet forms the inner limiting membrane. Lateral extensions from Müller cells contact neurons within cellular layers, synapses within plexiform layers, axons within the NFL, and blood vessels.
 Figure 2.5 Schematic illustration of neurotransmitters used by cells of the rod and cone pathways. The neurotransmitters are released within the outer and inner plexiform layers. The mGluR6 and AMPA receptors of the bipolar cells are also depicted. The former uses a G protein to control the intracellular concentration of cGMP that, in turn, regulates the Na+-Ca2+-cGMP-gated ion channel. |
Despite all of their connections, Müller cells are not part of the direct pathway for transfer of neural signals. Instead, they exert substantial influence over signal transmission by (1) maintaining the local extracellular environment needed for proper neuronal function, particularly with respect to the extracellular concentration of potassium; (2) adjusting concentrations of extracellular neurotransmitters; and (3) possibly buffering pH levels.
Astrocytes are found only on the vitread side of the retina, within the nerve fiber and ganglion cell layers. Two morphologic types of astrocytes develop from stem cells in the optic nerve. Each cell type preferentially contacts either nerve fiber bundles or blood vessels. Oligodendrocytes are not present in the primate retina, which is consistent with the absence of myelin in these retinas. However, the observation of myelinated nerve fibers in the retinas of about 0.5% of normal persons suggests that such a migration can occur during, and even after, development (see Chapter 3).
The inner plexiform layer is the zone of synaptic connection of bipolar and amacrine cells to ganglion cells and is much thicker than the OPL (Fig. 2.1). The IPL can be divided into sublaminae a and b. Sublamina a is the more proximal layer. It is
nearer the inner nuclear layer and is the zone within which bipolar, amacrine, and ganglion cells of the OFF-center pathway synapse. Sublamina b is the more distal layer. It is nearer the RGC layer and is the site of synaptic connections for the ON-center cell pathway.
nearer the inner nuclear layer and is the zone within which bipolar, amacrine, and ganglion cells of the OFF-center pathway synapse. Sublamina b is the more distal layer. It is nearer the RGC layer and is the site of synaptic connections for the ON-center cell pathway.
The retinal ganglion cell layer is, in fact, composed not only of RGCs, but also of “displaced” amacrine cells, astrocytes, endothelial cells, and pericytes (Fig. 2.1). The human retina contains about 1.2 million RGCs, although there is a very wide interindividual variability. It has been calculated that 69% of the RGCs in the human retina subserve the central 30 degrees of the visual field and are located adjacent to the fovea.
Dendrites of RGCs branch in well-defined strata within the IPL, and the specific region of stratification varies with the particular cell type. ON-center and OFF-center cells have dendrites that terminate in sublamina b or a of the IPL, respectively. RGC axons terminate in either the magnocellular or parvocellular layers of the lateral geniculate nucleus (LGN), and this also depends on the cell type. In the primate retina, there may be as many as 10 or more subpopulations of RGCs.
RGCs can be subdivided into two main types based on cell size. About 80% of the cells are small cells that are called “beta” or “P” cells, for parvocellular cells. These cells, as their name implies, synapse in the parvocellular layers of the LGN. From 5% to 10% of primate RGCs are large cells that are called “alpha” or “M” cells, for magnocellular cells. The axons of these cells synapse in the magnocellular layers of the LGN. Together, the P and M RGCs comprise 85% to 90% of all primate RGCs. The remainder of the ganglion cells is present in small numbers, is less well studied, and has functions other than for visual perception, such as the pupillary light reflex (see below) and the retinohypothalamic pathway, the latter believed to be an integral part of the pathway that regulates circadian functions.
From a clinical standpoint, the P and M cells have great significance. The P cells are color opponent (i.e., the center of the receptive field is maximally responsive to either red or green light, whereas the inverse is true for the surround), have small receptive fields, and have low sensitivity to contrast in the visual scene. Color opponency is generated by connections at both the OPL and IPL. The receptive field center of P ganglion cells near the fovea approximately equals the diameter of a single cone. These cells have linear response characteristics, meaning that their firing rate is proportional to the degree to which the center of the receptive field is stimulated or inhibited. Cells with this response property are also called “X” cells. Given these characteristics, the P cells are probably the major neural input to the optic nerve for visual functions such as central (i.e., Snellen) acuity, color vision, and fine stereopsis.
The M cells are spectrally “broadband.” Because of the combined input to these cells from all three cone types, their peak responsiveness at a given intensity of light is not determined by the wavelength. M cells have much larger receptive fields and are more responsive to luminance contrast than P cells. The majority of M cells have nonlinear responses to light. These cells typically show spikes of activity in response to changes in the visual scene, rather than showing activity that is proportional to the intensity of light and that matches the duration of light stimulation, as is the case with P cells. M cells with this physiologic profile are also called “Y” cells, to distinguish their responses from the X-like response profile of P cells. Lesions of the magnocellular pathway in monkeys impair high-temporal-frequency flicker and motion perception.
Most RGCs have several physiologic properties in common. First, their receptive fields have a concentric and antagonistic center-surround organization. This property probably derives at least in part from the effect of the lateral inhibition of horizontal and, possibly, amacrine, cells. Second, most respond either in an on or off manner to change in the luminance contrast or spectral distribution of light within their receptive field centers. This property is reflected in the location of dendritic ramification within the IPL. ON-center bipolar and ganglion cells ramify in sublamina b of the IPL, whereas OFF-center bipolar and ganglion cells ramify in sublamina a. Third, the responses to light of most RGCs are either sustained or transient, depending on the pattern of interconnections with amacrine cells within the IPL. Fourth, RGCs with sustained responses also show linearity of spatial summation; that is, the magnitude of their response is proportional to the degree of illumination within the center of the receptive field. In contrast, transiently responding RGCs briefly increase or decrease their firing rates in response to a change in illumination. Finally, RGCs maintain a continuous spontaneous discharge. This activity requires considerable energy but presumably has the advantage of allowing the cell to reflect more precisely the degree of illumination by either increasing or decreasing its firing rate.
As noted above, not all RGCs are P or M cells. Melanopsin (also called opsin 4, Opn4) is a photopigment that was first identified in frog skin and thereafter was detected in a small subpopulation of RGCs in vertebrates. These cells depolarize in response to light stimulation without any synaptic input from rods and cones; hence, their designation as intrinsically photosensitive RGCs (ipRGCs). The ipRGCs combine their direct, melanopsin-triggered photoresponses with signals derived from rods and cones and project to several targets in the diencephalon and midbrain involved in circadian rhythms and pupil responses to light (Fig. 2.6).
The ipRGCs constitute a small percentage of ganglion cells; in each human eye, about 3,000 of 1.5 million (0.2%) RGCs stain positively for melanopsin;
these cells are concentrated in the parafoveal region and at the far end of the nasal hemiretina. Like all RGCs, ipRGCs utilize the excitatory amino acid L-glutamate as their primary neurotransmitter; they also express pituitary adenylate cyclase-activating polypeptide (PACAP), which acts as a cotransmitter of glutamate in the retinohypothalamic pathway. There are several subtypes of ipRGCs that differ in their dendritic distribution in the IPL, melanopsin expression, physiologic responses to light, and connections. The melanopsin-containing ipRGCs are relatively spared in inherited mitochondrial optic neuropathies, may be selectively affected in glaucoma and may trigger photophobia in patients with migraine.
these cells are concentrated in the parafoveal region and at the far end of the nasal hemiretina. Like all RGCs, ipRGCs utilize the excitatory amino acid L-glutamate as their primary neurotransmitter; they also express pituitary adenylate cyclase-activating polypeptide (PACAP), which acts as a cotransmitter of glutamate in the retinohypothalamic pathway. There are several subtypes of ipRGCs that differ in their dendritic distribution in the IPL, melanopsin expression, physiologic responses to light, and connections. The melanopsin-containing ipRGCs are relatively spared in inherited mitochondrial optic neuropathies, may be selectively affected in glaucoma and may trigger photophobia in patients with migraine.
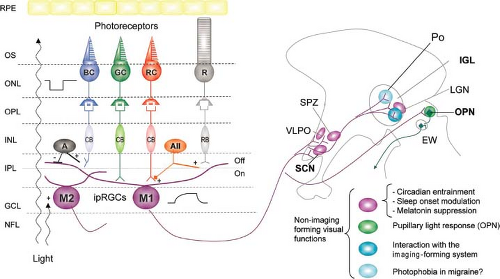 Figure 2.6 The melanopsin-containing intrinsically photosensitive retinal ganglion cells (ipRGCs) have a giant soma and long, sparsely branching dendritic processes that extend either into the outer (off) sublayer of the inner plexiform layer (M1 cells) or the inner (on) sublayer (M2 cells). These ipRGCs integrate their direct light responses with signals from rods and cones for non–imaging-forming functions. Rods provide excitatory inputs to ipRGCs sequentially via rod bipolar (RB), type II amacrine (AII), and cone bipolar (CB) cells. Red cones (RC, long wavelength, L-cones) and green cones (medium wavelength, M-cones) provide excitatory inputs to the proximal dendrites of ipRGCs through cone bipolar cells. In contrast, blue cones (BC, short wavelength, S-cones) trigger provide inputs presumably through cone bipolar cells and inhibitory amacrine (A) cells. The axons of ipRGCs exit the retina via the optic nerve and project to the suprachiasmatic nucleus (SCN), the subparaventricular zone (SPZ), the ventrolateral preoptic area (VLPO), and the intergeniculate leaflet (IGL) of the lateral geniculate nucleus (LGN), that are involved in circadian regulation, and to the olivary pretectal nucleus (OPN), which is a relay of the pupillary light reflex. Projections to the dorsal LGN provide an interface with the imaging-forming system; projections to dural-sensitive neurons in the posterior hypothalamus (Po) may contribute to light-induced exacerbation of migraine. EW, Edinger–Westphal nucleus; GCL, ganglion cell layer; INL, inner nuclear layer; IPL, inner plexiform layer; NFL, nerve fiber layer; ONL, outer nuclear layer; OPL, outer plexiform layer; OS, outer segment; RPE, retinal pigment epithelium. (Reprinted from Benarroch EE. The melanopsin system. Phototransduction, projections, functions, and clinical implications. Neurology. 2011;76:1422–1427, with permission.) |
The axon of an RGC is relatively long, and the body of the cell is the sole site for production of the components needed to maintain the health of the axon. The axon is a dynamic structure that requires nearly constant repair of its membranes, a process that is partly achieved by transport of proteins, enzymes, and other subcellular components (including mitochondria) to, and detritus from, sites up to the synapse.
Axon transport is bidirectional and simultaneous, with velocities that can be divided broadly into fast (i.e., hundreds of mm/day) and slow (<10 mm/day). Slow anterograde transport, which largely carries components that remain within the cell such as cytoskeletal proteins, constitutes the bulk (about 85%) of all movement within the axon. At least five different classes of proteins are transported at different velocities within RGCs, and these relationships vary during development. Fast anterograde transport is used to
carry neurotransmitter-containing vesicles. Retrograde transport, which is less well understood, proceeds at roughly half the maximum velocity of anterograde transport. It returns to the cell body substances that have been taken into the cell at its terminal synapse. Axon transport, whether anterograde or retrograde, is highly energy dependent, and the adenosine triphosphate (ATP) needed to sustain it is supplied by mitochondria that are distributed along the entire length of the axon.
carry neurotransmitter-containing vesicles. Retrograde transport, which is less well understood, proceeds at roughly half the maximum velocity of anterograde transport. It returns to the cell body substances that have been taken into the cell at its terminal synapse. Axon transport, whether anterograde or retrograde, is highly energy dependent, and the adenosine triphosphate (ATP) needed to sustain it is supplied by mitochondria that are distributed along the entire length of the axon.
The final common defect in pathologic swelling of the optic disc from almost any etiology is disruption of anterograde axonal transport of the RGC at the lamina cribrosa (see Chapter 4). The degree to which fast versus slow transport is disrupted probably varies according to the etiology.
Axons of RGCs synapse in the LGN, mesencephalon, pretectum, or one of several nuclei in the hypothalamus. The specific site of termination of the axon is related to the anatomy of the ganglion cell body. Generally, P- and M-cell axons synapse in the parvocellular and magnocellular layers, respectively. ON-center and OFF-center ganglion cells appear to remain functionally separated at the level of the LGN, although both channels converge at the visual cortex.
The nerve fiber layer of the retina is composed of axons of RGCs, astrocytes, components of Müller cells, and a very small number of efferent fibers to the retina, the functions of which are unknown (Fig. 2.1). The NFL is thinnest in the peripheral retina and thickest adjacent to the superior and inferior margins of the optic disc, where it measures about 200 μm in humans. The temporal and nasal NFL adjacent to the optic disc is about 1/10 as thick.
The gross organization of the NFL is characterized by three main features. The first is the papillomacular bundle, which contains nerve fibers originating from RGCs in the foveal area. Papillomacular fibers from RGCs on the nasal side of the fovea project directly to the optic disc, whereas those from RGCs on the temporal side of the fovea arch around the nasal fibers en route to the optic nerve (Fig. 2.7). The relatively early formation of the central retina relative to the peripheral retina gives rise to the second feature: the arching of axons from midperipheral and peripheral ganglion cells temporal to the fovea around the previously formed papillomacular bundle. A watershed line called the temporal raphe is present on the temporal side of the fovea, and this horizontal demarcation separates axons in the superior temporal retina from those in the inferior temporal retina. The raphe creates not only an anatomic separation, but also a physiologic separation between the superior and inferior regions of the temporal retina. The third anatomic feature of the NFL is the radial distribution of nerve fibers that enter the nasal aspect of the optic disc.
 Get Clinical Tree app for offline access 
|


