FIGURE 8.1 The retinas of vertebrates develop as evaginations from the anterior neural tube. The optic vesicles become patterned into domains of retinal pigment epithelium (RPE, gray), neural retina (blue), and the optic stalk connecting the developing retinal to the brain. In the next stage, a distinct optic cup forms as the retina folds into the RPE to become a twolayered epithelium; the most distal parts, near the rim of the cup, give rise to the ciliary epithelium, a two-layered epithelium, with an outer pigmented layer and an inner nonpigmented layer (red). The embryonic eye continues to grow, and even more distally than the ciliary epithelium, the iris develops at the rim of the cup, initially as a two-layered epithelium as well (green). (Modified from Principles of Regenerative Medicine, Atala A. et al., eds., Chapter 23 (T.R.), Regenerative Medicine for Diseases of the Retina, p. 419, Copyright Elsevier (2008)).
During gastrulation, neural induction transforms a region of ectoderm into the neural plate that contains the precursors of the central nervous system (CNS). The specification of the different regions of the CNS occurs around the same time, and the current understanding supports a model in which the same factors that induce neural tissue bias the cells to an anterior identity, telencephalon and diencephalon. More posterior regions of the CNS are induced by a second set of factors, sometimes called “transformers,” that promote more posterior fates. Indelibly labeling the cells in various regions of the neural plate in amphibians with vital dyes first allowed embryologists to fate map the regions of the neural plate, and along with transplant studies, they were able to show that the cells of the neural plate are specified very early to give rise to different parts of the CNS. The region of the neural plate that is specified to produce the optic cups and their derivatives is called the eye field. The eye field forms as a continuous domain at its anterior end of the neural plate but is soon split into two domains by Sonic hedgehog (Shh), a factor released by cells of the subjacent prechordal mesoderm. Mice lacking the Shh gene develop cyclopia, since the eye field is not repressed at the midline (4). In humans, mutations in the Shh gene cause congenital holoprosencephaly and ocular defects, including cyclopia (4,5).
The eye field is specified to produce the optic cup by its expression of a group of transcription factors, known as the eye-field transcription factors (EFTFs). These proteins bind to DNA and activate genes necessary for eye development (6). The first of these EFTFs to be identified was Pax6, a member of the paired class of homeodomain proteins; transcription factors of this family are involved in specifying the different regions of the embryo, including the neural tube. The eye-field cells express Pax6, as do the cells of the optic vesicle and the cells in the developing lens. Pax6 deletion in mice causes a failure of eye development around the optic vesicle stage; Pax6 mutations in humans cause a range of eye defects, including microphthalmia, Peters anomaly, foveal hypoplasia, and aniridia in humans (7). A homologous gene called eyeless was identified in Drosophila and is required for eye development in flies (8): misexpression of eyeless in other regions of the developing embryo induces ectopic eyes in various regions of the fly, like the leg or antenna (9). The fact that Pax6/eyeless has a conserved role in eye development across millions of years of evolution has led to the idea that Pax6/eyeless is a master control gene for eye development (10).
Although Pax6 is clearly an important factor in eye development, it is only one of many homeodomain transcription factors necessary for normal eye development. The coordinated actions of many genes together define the eye-field identity. There is evidence for EFTFs acting as complexes and also as a cross-regulatory pathway. Rx, a paired-type homeodomain transcription factor, is required at a very early stage in eye development; in mice, deletion of Rx nearly fully prevents eye development (11). Overexpression of Rx induces Pax6 expression, which subsequently induces expression of many other EFTFs. Experimental overexpression of each of the EFTFs has been done, and nearly all can induce ectopic eye–like tissue; however, coordinated overexpression of all of them (Pax6, Rx1, Six3, Six6, Lhx2, Nr2e1, and Tbx3) is sufficient to induce ectopic eyes in amphibia, even in nonneural areas of the frog. These experiments in frogs, together with similar studies in mice, support the hypothesis that the EFTFs form a cross-regulatory, feed-forward loop (6). Since similar genes are needed for eye formation in Drosophila and vertebrates, investigators have suggested that a common ancestor used some similar pathway (10). Although there are many differences between the compound eyes of arthropods and the camera-type eyes of vertebrates, the conservation of the genes that control development of photosensitive organs is striking.
PATTERNING THE DOMAINS OF THE OPTIC CUP
The next stage in eye formation in vertebrates requires several inductive interactions for proper development. The neural retina and the retinal pigment epithelium (RPE) both originate from the optic vesicles, but they ultimately become quite distinct; the neural retina is a multilayered structure with neurons and light responsive photoreceptors, while the RPE is a single epithelial layer of nonneural, cuboidal cells. The tissues surrounding the optic vesicle, as well as domains of the neuroepithelium immediately adjacent to the vesicles, play important roles in the patterning of the retina and RPE, as well as the ciliary epithelium and iris. If the optic vesicle is experimentally isolated from the surrounding tissues, the optic vesicles fail to form complex eyes and instead develop primarily as neural retina. Moreover, transplantation experiments have shown that if an optic vesicle is placed in a position in an embryo adjacent to the developing hindbrain, a second neural retina is formed from the presumptive RPE layer.
In addition to their morphologic differences, the two main domains of the optic vesicle, the RPE and the neural retina, can be distinguished from a very early stage by differences in their expression of distinct transcription factors. For example, Vsx2/Chx10 is expressed specifically in the neural retina, whereas a different transcription factor, Mitf, defines the domain of the presumptive RPE. These transcription factors play an important role in the specification of these two distinct tissues: loss-of-function mutations in Mitf cause the RPE to be respecified into neural retina, forming a duplicated retina; by contrast, mutations in Vsx2/Chx10 cause the neural retina to express RPE genes (for review, see (12)). Other transcription factors are also critical for RPE and neural retinal development. Otx2 is expressed in the RPE, and misexpression of Otx2 to the neural retinal domain, along with the signaling molecule Wnt, converts the presumptive neural retina into a second RPE layer (13). The development of the RPE is also regulated by signaling molecules. The mesenchymal (neural crest–derived) tissue that surrounds the optic vesicle promotes RPE-specific gene expression and represses neural retinal genes. The addition of the signaling factor, activin, to cultures of chick embryo optic vesicles can substitute for the extraocular tissue to promote RPE (14) as can other members of the TGF-beta superfamily (15). In mice, targeted deletion of the Bmp7 gene disrupts ocular development, though the phenotype is more severe than just failure of RPE development, and other evidence indicates that bone morphogenic protein (BMPs) have multiple roles in eye formation. Shh is also involved in the specification of the RPE domain, specifically in the ventral optic vesicle; inhibition of Shh in frog embryos causes defects in ventral RPE formation, while inhibition of Shh signaling in mice blocks ventral RPE development (16).
The specification and growth of the neural retina domain also requires several signaling molecules. Fibroblast growth factors (FGFs) are important for proper development of the neural retina. Several different FGFs are expressed in the lens ectoderm and the neural retinal domain (17,18). Overexpression and inhibition experiments in zebra fish, chick, and mouse embryos support key roles of FGFs in patterning the neural retinal domain. For example, the addition of exogenous FGF to chick embryo optic vesicle cultures induces the presumptive RPE layer to become a second neural retina (18), while blocking FGFs with antibodies in similar cultures prevents neural retina formation. Inhibition of FGF signaling in mice by conditional deletion of Shp2 (a necessary component for FGF signal transduction) causes the affected region of the neural retina to instead develop as RPE (19). The model of ocular patterning that has emerged from these, and other studies, is as follows: FGF promotes neural retinal specification in the optic cup, in part by inducing Vsx2. Vsx2 in turn represses Mitf in the retinal domain. The presumptive RPE is then specified by several signaling molecules, including Shh, Wnt, and activin/BMP, some of which are derived from the surrounding mesenchyme (activin, Indian hedgehog) and some of which (e.g., Sonic hedgehog, BMP7) come from within the neuroepithelium, to promote Mitf, which in turn represses Vsx2 (12).
HISTOGENESIS OF THE RETINAL CELL TYPES
After the primary domains of the optic vesicle have been specified, their growth accelerates, particularly in the neural retina. The neurons and Müller glial cells of the retina are generated by progenitor cells through multiple mitotic cell divisions. These progenitors divide rapidly and, in humans, produce hundreds of millions of cells between the 10th and the 24th gestational week. In the early part of retinal development, there is a nearly exponential increase in cell numbers, reflecting the symmetric cell divisions of the progenitors; however, some of the cell divisions also produce postmitotic neurons, even at very early stages of retinal histogenesis. The different types of cells in the retina are produced by the progenitor cells in a regular sequence. The regular “birth order” of retinal cells was first described by Richard Sidman, using the 3H-thymidine “birthdating” technique (Fig. 8.2A,B) (20). The production of the different types of retinal cells can be divided into two phases. In the early phase, most of the ganglion cells, cones, and horizontal cells are produced, whereas most of the rod photoreceptors, the bipolar cells, and the Müller glial cells are produced in a second phase (21,22). Amacrine cells do not fall as neatly into one or the other phase. Although there is clear regularity in histogenesis in the retina, there is also a central-to-peripheral gradient, such that peripheral retina may still be producing ganglion cells while central retina is producing later cell types.
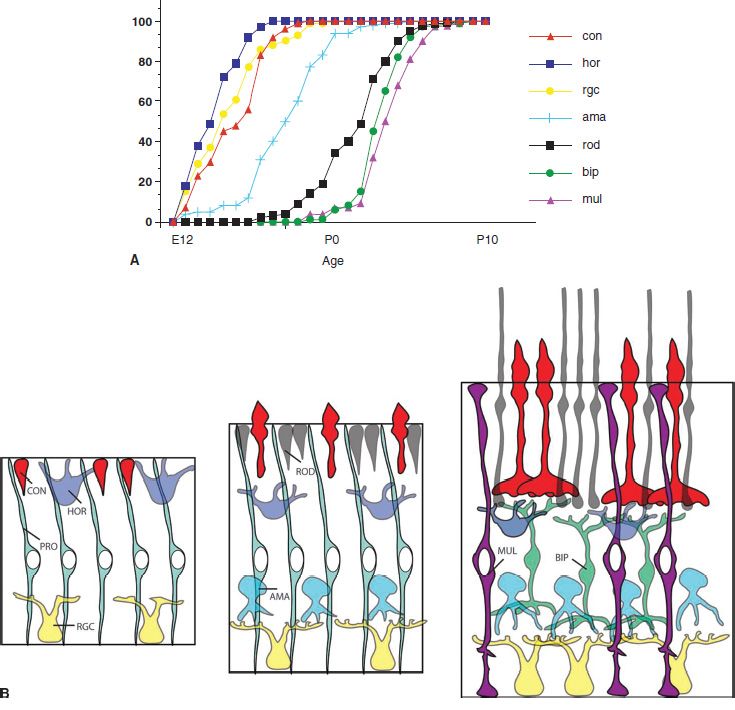
FIGURE 8.2 Each retinal cell type is generated over a slightly different time course during retinal histogenesis. The graph at the top shows the running total of each cell type as a function of age in the rat. The lower figure shows schematically the addition and integration of new cells during the histogenesis of the retina at three different ages, corresponding to the chart of birthdating above. (A: Modified from La Vail MM, Rapaport DH, Rakic P. Cytogenesis in the monkey retina. J Comp Neurol 1991;309(1): 86–114).
The progenitor cells of the retina are for the most part able to generate all the different types of cells and, therefore, have been called multipotent progenitors (23,24). The multipotential nature of these cells has been demonstrated by tracking the lineages of individual cells, after infection of progenitors with a retrovirus containing a reporter gene (23,24) or following injection of the cells with an impermeant dye (25,26). More recently, it has been possible to track the lineages of progenitors that express specific transcription factors, using inducible cre-recombinase methods (27,28). These studies have shown that progenitors can produce multiple different types of retinal neurons and Müller glia up to their last cell division; however, there do appear to be at least three distinct types of progenitors in the mouse: (i) progenitors that express Ascl1 can produce moderate-sized clones with all types of retinal neurons, except ganglion cells (Fig. 8.3); (ii) Neurog2-expressing progenitor cells produce clones with no more than two cells and are likely in their last cell division (nevertheless, they can also make ganglion cells); and (iii) true multipotent progenitor cells that can make large clones containing all cell types (27).
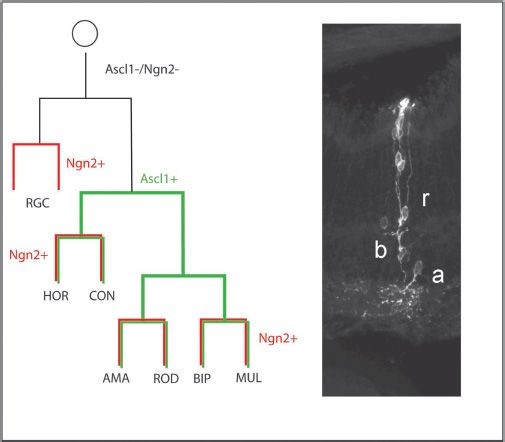
FIGURE 8.3 Tracking the lineages of retinal cells shows that there are several types of multipotent progenitor cells that underlie cellular diversity. The schematic diagram shows examples of lineages of (1) a retinal progenitor that expresses Ascl1 (green) that can produce a clone of cells that include all cell types (HORizontal cells, CONes, AMAcrine cells, BIPolar cells, ROD photoreceptor cells and MULler glia), except ganglion cells, which are produced by (2) an earlier progenitor (black) that is competent to produce ganglion cells, and then produce an Ascl1+ progenitor. The diagram also shows the results of tracking the Neurog2-expressing progenitor lineage in red. Neurog2 is expressed in both the Ascl1+ and Ascl1− progenitors but only at the last division of the progenitor cells.
The fact that the different types of retinal cells are generated in sequence has led to several hypotheses about the nature of the mechanisms that underlie this phenomenon. One proposal is that each cell type induces the progenitor cells to produce the cell type that follows in the sequence. In this model, the first cell type generated by the progenitor cells, the retinal ganglion cells, produces a signal to the progenitor cells to direct them to begin producing horizontal cells, the next cell type in the sequence; the horizontal cells in turn signal to the progenitors to instruct them to make cone photoreceptors, and so on. Cell culture experiments provide some support for this type of mechanism; progenitor cells isolated from the early stages of retinal development predominantly differentiate into retinal ganglion cells (29), whereas culturing the progenitor cells with retinal cells from later stages of development biases them to adopt later cell identities (30–32). In other words, the progenitors can be influenced by their surrounding microenvironment to form the types of cells they produce. One of these factors is the signaling molecule Shh, mentioned earlier in the chapter in the context of the eye field. Shh is expressed by the retinal ganglion cells, regulates the rate of proliferation and timing of cell cycle exit of the progenitor cells, and consequently determines the number of ganglion cells in the retina (33).
An alternative model to explain the sequential production of the different retinal cell types proposes that the progenitors undergo a progressive change in their “internal state,” like a clock ticking through the different cell fates (34,35). In this model, the progenitor cells progressively change in their competence to produce different types of cells; their initial state has a bias for producing retinal ganglion cells, but after a day some fraction of the progenitor cells shift their competence to become biased to produce later cell types, for example, horizontal cells, followed by cone photoreceptors, and so on. The molecular mechanism underlying such a progressive shift in competence could be a cascade of transcription factors, and Ikaros genes have been shown to be involved in this process (36). Recent data have also implicated miRNAs in regulating the developmental timing. Deletion of the Dicer gene, which is necessary for miRNA production, leads to a failure in the progenitor cells to progress beyond the early state, and they continue to generate early cell fates, like ganglion cells, cones, and horizontal cells for the entire period of retinogenesis (Fig. 8.4) (37,38).
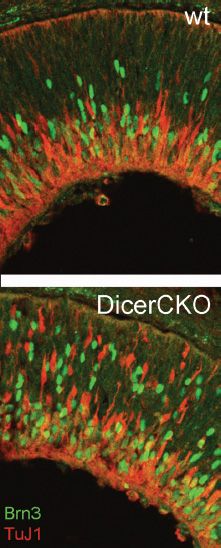
FIGURE 8.4 MicroRNAs are required during retinal development to allow the progenitors to make late retinal cell types. When the Dicer gene is knocked out in mouse retinas, the progenitors overproduce the early-generated ganglion cells (Brn3+) and do not progress to the state where they can generate late cell types like rods, bipolar cells, and Muller glia.
The mechanisms by which the progenitors progressively change in their competence are not understood, but quite a bit is known about the transcription factors that are necessary for the differentiation of different types of retinal cells. One transcription factor discussed earlier in the chapter as important in the eye field, Pax6, is also critical for cell fate determination later in retinal development. Conditional deletion of Pax6 in retinal progenitors causes a dramatic restriction in their competence such that they only produce amacrine cells (39). Conversely, deletion of the transcription factor FoxN4 produces the complementary result, and in these mutants progenitor cells produce all cell types except amacrine cells (40). Other transcription factors expressed by progenitors also play important roles in the production of cell diversity in the developing retina. For example, deletion of the gene Atoh7 (Math5) causes a dramatic reduction in ganglion cell production (41), whereas conditional deletion of Otx2, another EFTF, leads to a loss of bipolar and photoreceptor cells (42–44); the transcription factors Ptf1a, NeuroD1, and Math3, expressed in amacrine cells, are necessary for amacrine cell development (40); the Nrl transcription factor is required for rod photoreceptor development, and loss of this gene causes the progenitor cells to produce as short-wavelength-sensitive cones instead (45); mutations in the Chx10/Vsx2 gene result in an absence of nearly all bipolar cells (46); conditional deletion of the Prdm1/Blimp transcription factor switches the photoreceptors to bipolar cells (47); and deletion of Ascl1, a bHLH transcription factor, results in an increase in the Müller glia and a concomitant reduction in late-born retinal neurons: bipolar cells and rods (34,48,49). The way in which these transcription factors interact remains a significant challenge for the field.
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


