FIGURE 6.1. The cyclooxygenase cascade. COX-1 and COX-2 catalyze the biosynthesis of prostanoids (prostaglandins and thromboxane) from membrane-derived arachidonic acid. The prostanoids then bind to their cognate GPCRs (G-protein coupled receptors) on the surface of target cells, initiating and affecting a wide range of physiological and pathological processes. Some of the ocular effects are listed.
Selective inhibition of COX-2 prevents pathological NV in the cornea, retina, and choroid. Two investigative teams have used rodent models of cautery-induced corneal NV to demonstrate that indomethacin, a nonselective COX inhibitor, and NS-398, a COX-2–selective inhibitor, significantly inhibit corneal NV and the concomitant model-induced increase in corneal PGE2 and VEGF.28,29 These findings suggest that COX-2 and the PGs mediate corneal NV in a VEGF-dependent manner.
COX-2 has also been implicated in the retinal NV that characterizes PDR and ROP. Several investigators have used streptozotocin (STZ)-induced diabetic rats to study the effect of COX-2 inhibition on the development of retinopathy. Treating the animals with celecoxib, an NSAID with moderate COX-2 selectivity, led to significant reductions in retinal PGE2, VEGF mRNA expression, and vascular leakage.30–32 Treating diabetic rats with topical nepafenac, a nonselective COX inhibitor, has been shown to inhibit the functional abnormalities and retinal vasculopathy observed in rats with STZ-induced diabetes.33 These studies suggest a functional link between the COX pathway and PDR. Other investigators have used rodent models of ROP to define the involvement of COX-2 in retinal NV. Rodents treated with rofecoxib, etodolac, and APHS, COX-2 selective NSAIDs, and topical nepafenac demonstrate reduced retinal NV34–36 (Fig. 6.2). In the case of etodolac, this effect was abolished by the addition of exogenous PGE2, supporting a role for COX-derived PGs in mediating retinal NV. The same study provides data suggesting that the effect of PGE2 is mediated through the EP3 receptor. Signaling through EP3 downregulates the antiangiogenic factor thrombospondin-1 (TSP-1) and its receptor, CD36. Another study demonstrates that the EP4 receptor is involved in mediating angiogenic cell behaviors (VEGF production and endothelial cell proliferation and tube formation) and experimentally induced CNV and ROP using in vivo animal models.37
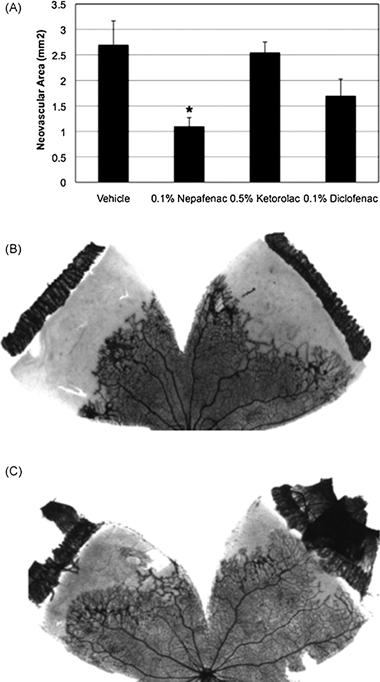
FIGURE 6.2. The effect of topical nepafenac, ketorolac, and diclofenac on the severity of OIR in the rat. A: Drugs were administered topically, QID, from P14-P19. Nepafenac significantly reduced (*p ≤ 0.007) OIR-induced retinal NV. Ketorolac and diclofenac failed to demonstrate an effect. Each bar represents the mean ± SEM. B: A representative image of NV in vehicle-treated eyes. C: A representative image of NV in nepafenac-treated eyes. As demonstrated by representative ADPase-stained retinal flat mounts, nepafenac significantly reduced retinal NV. (Reproduced from Yanni et al. Brain Res Bull. 2010;81:310, with permission.)
In rodent models of CNV, etodolac and lumiracoxib, NSAIDs with COX-2 selectivity, significantly inhibited CNV leakage, lesion size, and the development of choroidal neovascular membranes (CNVMs; angiogenic outgrowths that infiltrate the retina).38,39 Additionally, topical ocular administration of nepafenac inhibited laser-induced CNV.36 These studies suggest that COX-2 mediates various aspects of CNV.
In conclusion, ocular studies suggest that COX-2 and the PGs play a role in mediating NV, involving at least some degree of interaction, in some subset of cases, with VEGF.
Angiopoietins
There are four angiopoietin ligands (Ang1 to 4) and two related receptors, Tie1 and Tie2.40 The best studied angiopoietins are angiopoietin 1 (Ang1) and angiopoietin 2 (Ang2). In vivo, Ang1 and Ang2 function in a complementary fashion to maintain vascular integrity. Ang1 promotes the integrity of vasculature by recruiting mural cells to blood vessels.41 Transgenic mice deficient in Ang1 exhibit dissociation of endothelial cells and mural cells, severely impairing vascular function and leading to embryonic death.42,43 Initially, Ang2 was identified as an Ang1 antagonist, inhibiting Tie2 activation by Ang1. Transgenic mice overexpressing Ang2 have phenotypes similar to those of Ang1-deficient mice, where the dissociation of endothelial cells and mural cells severely disrupts blood vessel formation in the embryos. In adult mice and humans, Ang2 is expressed only at sites of vascular remodeling.44 The primary role of Ang2 in angiogenesis appears to be in concert with VEGF. In the presence of VEGF, Ang2 destabilizes blood vessels by disrupting interactions between endothelial cells and mural cells, thus enhancing VEGF-induced endothelial cell proliferation and migration. In the absence of VEGF, however, Ang2 accelerates vessel regression.45 This relationship was demonstrated by a study of the hyaloid vasculature. In the presence of exogenous VEGF, Ang2 promoted a rapid increase in hyaloid capillary diameter, remodeling of the basal lamina, proliferation and migration of endothelial cells, and stimulated the sprouting of new blood vessels. When VEGF was inhibited, Ang2 induced endothelial cell apoptosis, resulting in hyaloid vessel regression.46
In contrast to Tie2 signaling, Tie1 signaling is still poorly understood. Experiments have shown that Tie1 activation can initiate the PI3K–AKT signaling pathway to promote cell survival.47 These findings suggest that Tie1 elicits signaling cascades similar to those initiated by Tie2. However, Tie1-specific ligands have not been identified, suggesting that Tie1 might be activated by other mechanisms. Recent work suggests that Tie1 is proteolytically cleaved following endothelial cell activation. The exposure of cultured endothelial cells to phorbol-12-myristate-13-acetate (PMA), VEGF, or shear stress induces cleavage of Tie1.48,49 The biological consequence of this process is not understood, but the truncated form of Tie1 that is generated is thought to contribute to Tie2 signal transduction. For a review of angiopoietin-mediated signal transduction cascades and bioactivity, see Figure 6.3.
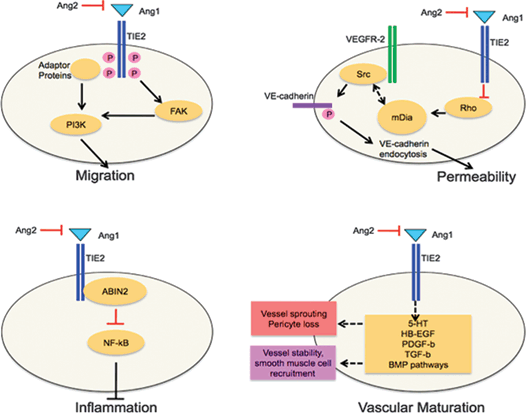
FIGURE 6.3. The effect of angiopoietins on endothelial cell (EC) signal transduction and angiogenic cell behaviors. Ang1-mediated phosphorylation of focal adhesion kinase (FAK) and other adaptor proteins leads to phosphoinositide 3-kinase (PI3K)–mediated cell migration. In the absence of Ang1, Src is not sequestered by mammalian diaphanous (mDia). VEGF stimulation activates Src, which leads to the internalization of VE-cadherin. This destabilizes interendothelial junctions and increases permeability. Tie2 activation leads to the recruitment of the intracellular protein ABIN2 (A20 binding inhibitor of NF kappa B2). This blocks the nuclear factor-kappa B (NF-κB) pathway, therefore exerting anti-inflammatory effects. Ang2 counteracts this response, leading to inflammation. Ang1–Tie2 signaling is required for vascular maturation (purple box). Ang2 is an antagonistic regulator of ECs and thereby leads to vessel destabilization and pericyte dropout (red box). The exact molecular mechanisms of this process are poorly understood. The possible regulatory molecules that are involved in this process are listed. (Adapted from Augustin HG, Koh GY, Thurston G, et al. Control of vascular morphogenesis and homeostasis through the angiopoietin-Tie system. Nature Rev Mol Cell Biol. 2009;10:165–177 with permission.)
Particularly important in the context of ocular disease is the effect of hypoxia on angiopoietins. Oh and coworkers have shown that Ang1 and Tie2 are stable in microvascular endothelial cells exposed to hypoxia, whereas Ang2 expression increases.50 Additionally, Ang2 expression in endothelial cells is tightly controlled by several angiogenic inducers including VEGF, FGF-2, and TNF-α and by environmental changes including high glucose.50–54 In vivo, Ang2 expression is greatly increased in the neovascular growth accompanying OIR in mice.50 These findings suggest that Ang2, acting in concert with VEGF and other angiogenic inducers, contributes to the ocular NV characterizing several sight-threatening diseases. Consequently, several methods of Ang2 inhibition are currently under assessment for inhibition of tumor growth and retinal NV in preclinical tests.55,56
Erythropoietin
Apart from its essential role in the regulation of red blood cell production, Erythropoietin (Epo) is involved in diverse nonhematopoietic biological functions. Various types of VE cells express the Epo receptor (EpoR), and Epo promotes the migration and proliferation of endothelial cells in different in vitro experimental models.57–60 During development, Epo–EpoR signaling is required not only for fetal liver erythropoiesis but also for physiologic angiogenesis in the developing mouse embryo.61,62 A series of studies suggests that endogenous Epo–EpoR signaling contributes to the angiogenic aspect of wound healing, a feature that is inhibited by neutralizing anti-Epo antibodies and recombinant soluble EpoR.63 In the central nervous system, where Epo is produced by astrocytes and EpoR is expressed by neurons, administration of soluble EpoR is associated with enhancement of ischemic brain injury in rats, demonstrating that Epo plays an important role in the brain’s response to neuronal injury.64 Epo and its novel derivatives have emerged as major tissue-protective cytokines that are being investigated in human studies involving both neurological and cardiovascular diseases.65
Although Epo is a survival factor for retinal photoreceptors,66 significant upregulation of endogenous Epo in the vitreous of patients with diabetes was found to be associated with PDR.67 Furthermore, Epo blockade was shown to inhibit retinal NV in the ischemic mouse retina, suggesting that Epo signaling may be a potential therapeutic target in the pathological angiogenesis of PDR.67 In a meta-analysis of Epo treatment for prematurity-related anemia, early Epo administration was associated with a significantly increased risk of ROP,68 an effect that may be associated with EpoR activation on endothelial cells and NV.69 This finding, however, was contradicted by those of Shah et al.,70 who found no significant difference in the onset or progression of ROP or the incidence of plus, prethreshold, or threshold disease between rhEPO-treated and nontreated groups of very low birth weight infants, all of whom received ROP treatment. In conclusion, although Epo-EpoR signaling has been implicated in various angiogenic processes, its precise role in ocular angiogenesis has yet to be elucidated.
Interleukins
The interleukins (IL) comprise a family of soluble proteins that mediate the interaction between immune cells and inflammatory cells. Today, at least 34 ILs and IL subtypes have been identified.
For some time, it has been known that the inflammatory response can solicit an angiogenic response. Because ILs induce the proliferation of immune cells, investigators hypothesized that some ILs might affect endothelial cells in the same way. In an early study, BenEzra et al.71 tested IL-1α, IL-1β, and IL-2 in a model of corneal angiogenesis. IL-1α and IL-1β induced a consistent angiogenic response. In another study, corneal explants exposed to IL-1β demonstrated an upregulation of IL-8, which has been shown to be proangiogenic.72
There is evidence that IL-4 may exert both proangiogenic and antiangiogenic effects, depending on the experimental setting. In the eye, IL-4 induced NV in the rabbit corneal micropocket assay.73
In human cerebral endothelial cells, IL-6 stimulates endothelial cell proliferation and migration in a dose-dependent manner and stimulates a concurrent increase in VEGF expression.74 Interestingly, experiments designed to test for possible signaling mechanisms revealed an IL-6– dependent activation of VEGFR-2 and MMP-9 overexpression. MMP-9 has been shown to be important in the development of retinal NV.75 Of particular clinical relevance, circulating blood endothelial progenitor cells (EPC) have received considerable attention because they home to sites of angiogenesis. IL-6 stimulates EPC proliferation, migration, and tube formation.76 These findings suggest that IL-6 has the potential to stimulate EPC involvement and incorporation into ocular neovasculature.
IL-8 has been shown to be proangiogenic. This effect may be due to the ability of IL-8 to inhibit apoptosis and upregulate MMP-2 and MMP-9.77 Like MMP-9, MMP-2 is involved in retinal angiogenesis.75
Experimental evidence suggests that IL-10 may exert a proangiogenic effect in the context of ocular angiogenesis. In a mouse model of OIR, IL-10–deficient mice showed less NV than did wild type controls.78 The authors attribute this response to the finding that wild type macrophages produced IL-10, which induced higher levels of retinal VEGF, contributing to the pathology. In another study of inflammation-induced corneal NV, IL-10– deficient mice demonstrated a delayed NV response.79 These studies suggest that IL-10, in the context of retinal and corneal NV, contributes to the angiogenic phenotype. IL-10 has also been hypothesized to contribute to the angiogenic phenotype in AMD. It is believed that during the pathogenesis of AMD, macrophages can switch from an antiangiogenic to a proangiogenic phenotype.80–82 Studies in mice have shown that low levels of IL-10 result in an antiangiogenic macrophage phenotype.82 According to theory, young macrophages express low levels of IL-10, leading to increased levels of IL-6 and IL-12, which have antiangiogenic activities in this context. Furthermore, this phenotype is characterized by high levels of FasL and TNF-α, proteins that mediate an antiangiogenic effect by inducing apoptosis in angiogenic endothelial cells. Increasing age induces a polarized phenotype that is proangiogenic, characterized by increased levels of IL-10, which is functionally linked to decreased levels of IL-6, IL-12, FasL, and TNF-α.81,82
IL-13 and IL-17 are also known to be involved in the angiogenic cascade in nonocular contexts.83–85 In the eye, however, their role(s) are not clear.
In summary, the ILs are a family of soluble factors with pleiotropic roles. In the eye, ILs have demonstrated both proangiogenic and antiangiogenic effects, which are dependent upon context and environment. This finding necessitates further research into the precise roles of specific ILs before they can be exploited as a targeted therapy for human use.
Other Growth Factors
Platelet-derived growth factor (PDGF) has been shown to promote cell survival, induce endothelial cell proliferation, and recruit pericytes to the retinal vasculature.86,87 Rats that were exposed to a model of OIR and treated with an inhibitor of the PDGF receptor demonstrated increased VEGF, pericyte dropout, and exacerbated angiogenesis.88 However, in another study, targeting PDGF-BB and VEGF in combination led to a greater reduction in corneal and choroidal NV than when either protein was targeted alone.89 These studies suggest that PDGF mediates context-specific aspects of ocular NV, both alone and in combination with VEGF.
Hepatocyte growth factor (HGF) induces the proliferation of hepatic and renal cells. Recently, a potential role of HGF in retinal angiogenesis has been identified.90,91 Levels of HGF are increased in the vitreous and subretinal space of patients with PDR and ROP.90,92,93 Transcriptome data suggest a possible synergy between VEGF and HGF. A three-way transcriptome comparison of HUVEC treated with VEGF, HGF, or VEGF + HGF was performed. The combination treatment revealed altered expression of a nonoverlapping set of genes that were unperturbed when these cells were treated with VEGF or HGF alone.94 These studies suggest that HGF promotes retinal NV, possibly demonstrating synergy with the proangiogenic effects of VEGF.
Placental growth factor (PlGF) is a structural and functional homologue of VEGF. However, in contrast to VEGF, PlGF only binds and activates VEGFR-1.95,96 PlGF is a weak mitogen for some endothelial cell types.95 Bovine retinal endothelial cells treated with PlGF and FGF-2 in combination demonstrated increased tube formation and secondary sprouting, suggesting a mechanism for its angiogenic influence.97
Transforming growth factor-beta (TGF-β) is generally an inhibitor of endothelial cell proliferation. Endothelial cells treated with TGF-β demonstrate increased expression of plasminogen activator inhibitor-1 (PAI-1), suggesting a possible mechanism for the antiangiogenic capacity of TGF-β.98,99
Tumor necrosis factor-alpha (TNF-α) inhibits endothelial cell proliferation, and administering high doses of TNF-α selectively destroys tumor vasculature.100,101 However, several in vivo studies indicate that TNF-α demonstrates a proangiogenic role.101 These studies indicate that TNF-α induces proangiogenic VEGF expression in various cell types, which may explain a mechanism for its angiogenic activity.102 In the eye, wild type and TNF-α–deficient mice were exposed to OIR. The TNF-α–deficient mice demonstrated increased intraretinal vascular development and significantly reduced preretinal NV in response to OIR exposure, suggesting that TNF-α is involved in promoting ocular angiogenesis.103
KEY POINTS
VEGF is the principal growth factor responsible for the pathological angiogenesis characterizing a number of ocular diseases. However, recent evidence suggests that several other factors play pivotal roles in the angiogenic cascade in the eye. Exploiting the therapeutic potential of these factors, alone or in combination with VEGF-directed therapies, may enhance the current standard of treatment for ocular angiogenic diseases.
- FGF-2 is thought to act in synergy with VEGF, promoting angiogenesis.
- COX-2 and its PG products are thought to play a proangiogenic role in ocular NV. Inhibiting COX activity with NSAIDs significantly reduces pathological angiogenesis in the eye.
- Ang2 acts in concert with VEGF to promote vessel destabilization and the angiogenic phenotype. Preclinical tests of Ang2 inhibition are currently underway.
- Epo-EpoR signaling has been implicated in a number of angiogenic processes and diseases. There are conflicting findings about the precise role of Epo in the eye.
- IL-1, IL-4, IL-6, IL-8, and IL-10 appear to promote ocular angiogenesis. Other members of the IL family, for example, IL-12, appear to be antiangiogenic. The literature suggests that the specific roles of each of the ILs are disease and context dependent. This may complicate development of IL-targeted treatment, but once developed, these drugs may provide a more directed therapeutic with fewer undesirable side effects.
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


