FIGURE 6.1 Cross-sections of 6.5 WG fetal choroid demonstrate hemovasculogenesis (A–F). Erythroblasts (bright pink cytoplasm) in the CC layer can form solid cord-like structures (double arrow) without a lumen (A). Erythroblasts and hematopoietic and vascular cells develop in situ with erythroblasts sometimes forming a lumen (arrow in D). Eventually, the outer cells become primarily ECs and the inner cells become primarily erythroblasts (F). Free erythroblasts (arrowheads in D–F) are present in stroma of choroid. The monolayer of RPE cells is visible at the top in (A–C). (Scale bar = 10 µm; Giemsa-stained JB4 sections.) (From Werner JS, Chalupa LM. The New Visual Neurosciences. Cambridge, MA: MIT Press, 2013, with permission.)
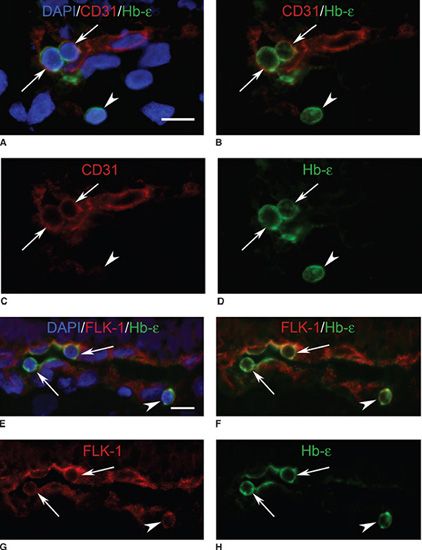
FIGURE 6.2 Colocalization of embryonic hemoglobin (Hb-ε) and endothelial markers (CD31 and VEGFR-2 or FLK-1) in developing CC. A–D: Hb-ε (green) and CD31 (red) are colocalized in cells of the developing CC (arrows) and single cells within the choroidal stroma (arrowhead). E–H: Hb-ε and FLK-1 (red) coexpression in cells lining a developing lumen (arrows) and in cells located outside of the structure (arrowheads). (Scale bars = 10µm; counterstained with DAPI, blue.) (From Lutty et al. Development of the human choriocapillaris. Eye [London] 2010;24:408–415.)
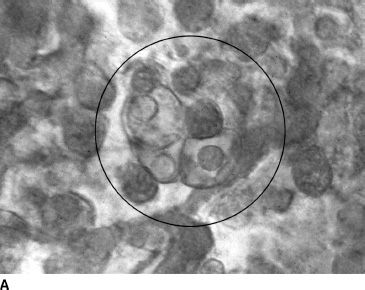
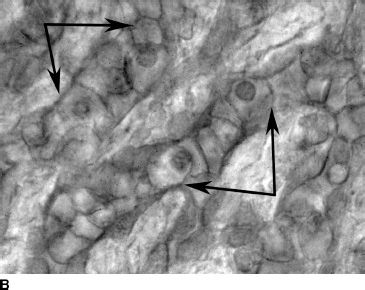
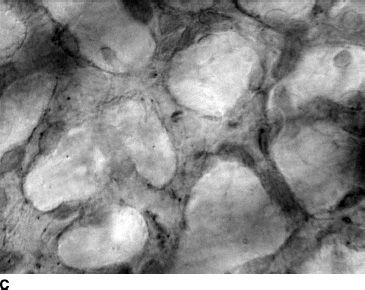
FIGURE 6.3 CD39-immunolabeled flat choroids showing the pattern of the developing CC in embryonic and fetal human eyes. At 5.5 WG (A), CD39-positive cells and erythroblasts are organized in blood island–like structures (circle). At 9 WG (B), highly cellular linear cord-like structures without apparent lumen have formed. By 12 WG (C), the capillaries are narrower, lumens have formed, and cellularity has decreased markedly.
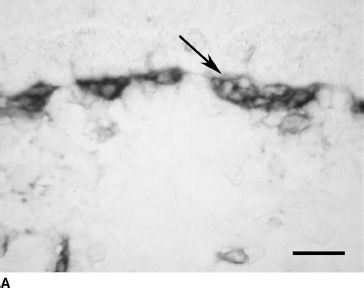
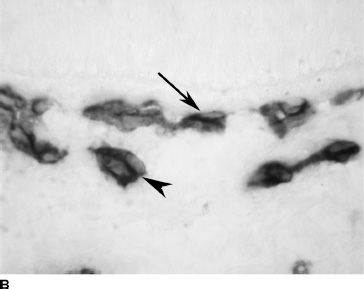
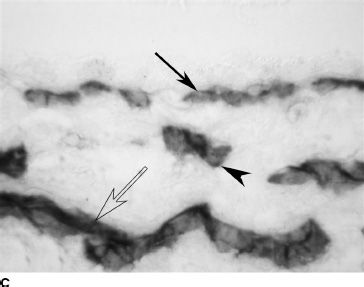
FIGURE 6.4 CD31 immunolabeling of choroidal sections from embryonic eyes at 6 WG (A) and fetal eyes at 12 WG (B) and 20 WG (C). Only a highly cellular rudimentary CC with poorly defined lumen (arrow) is present at 6 WG (A). At 12 WG (B), vessels are budding from the CC into the deeper choroid (arrowhead). At 22 WG (C), the CC (arrow), medium-size vessels of the Sattler layer (arrow head) and the larger outer blood vessels (open arrow) are all present. (Scale bar = 30 µm.)
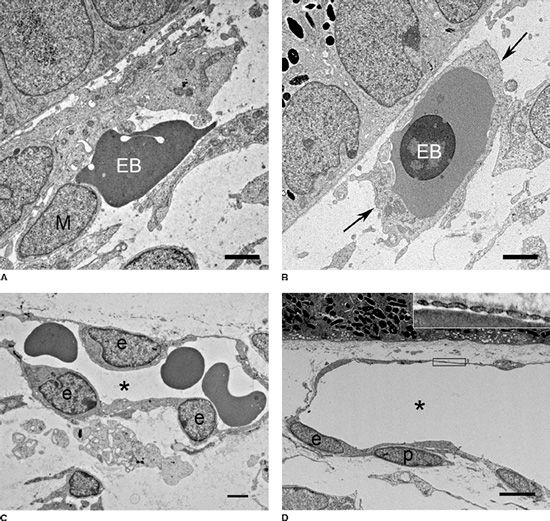
FIGURE 6.5 Ultrastructure of the developing embryonic and fetal human CC at 6.5 WG (A,B), 16 WG (C), and 22 WG (D). In ultrathin sections from 6.5 WG choroid (A,B), free progenitor cells (M) make contact with free erythroblasts (EB). In other areas, immature ECs envelop the erythroblasts to form blood island–like structures (B). The RPE is present in the upper left-hand corner in (A) and (B). By 16 WG, lumens are apparent (asterisk); the EC nuclei (e), with more organized chromatin, were reduced in volume and had decreased cytoplasmic projections. By 22 WG (D), lumens were broad and flat, ECs were thin, and fusiform and definitive pericytes (p) were present on the outer surface of the capillaries. Fenestrations were present along the inner aspect of the CC at this stage of development (inset) on the retinal side of CC lumens under RPE (top). (Scale bars = 20 µm [A,B]; 2 µm [C]; 4 µm [D].) (From Werner JS, Chalupa LM. The New Visual Neurosciences. Cambridge, MA: MIT Press, 2013, with permission.)
The functional properties of the CC were evaluated by performing immunohistochemistry of three enzymes known to be important in adult CC function: carbonic anhydrase IV (CA IV), endothelial nitric oxide synthase (eNOS), and alkaline phosphatase (APase) (15). CA IV, an ectoenzyme on ECs, controls local pH when coupled to the electrogenic sodium bicarbonate cotransporter (NBC1) (16–18). CA IV and eNOS were expressed at low levels as early as 8 WG, during hemovasculogenesis (5.5 to 9 WG) (11). eNOS’s function is presumably vasodilation of blood vessels but mostly slit-like lumens were present at 8 WG (11). Neuronal NOS (nNOS) on the other hand was present in the nuclei of vascular progenitors in CC, retina, and vitreous, as well as in the nucleus of RPE cells and scattered progenitors throughout the choroid (Fig. 6.6) (19). APase is a marker for choroidal blood vessels in adult human (15) and it was present in forming CC already at 7 WG, even though the primitive ECs were just starting to differentiate from hemangioblasts. However, the observation of high APase activity in the blood island–like formations is not unexpected, because it is used extensively as a marker for human embryonic stem cells and other progenitors (20,21).
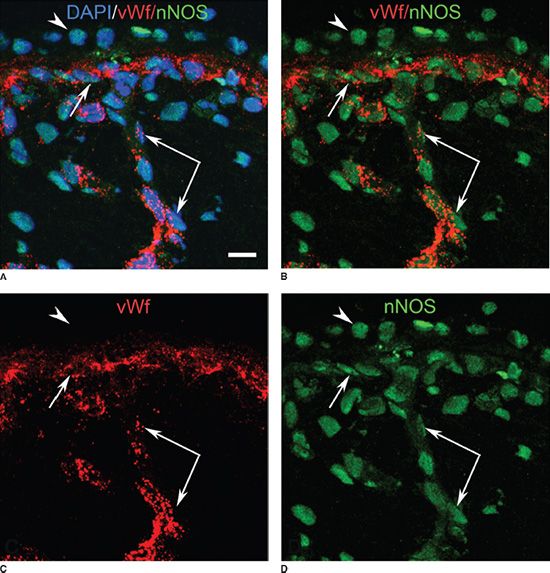
FIGURE 6.6 Choroid of a 21 WG fetal eye labeled for vWf (red) and nNOS (green) and DAPI (nuclei) (A) and shown as single (C,d) and multiple channel (A,B) images to demonstrate colabeling. Nuclear nNOS expression is seen in RPE (arrowhead), ECs of CC (arrow), and large choroidal vessels (paired arrow). Nuclei were counterstained with DAPI (blue). (Scale bar in [A] = 10 µm.) (From Werner JS, Chalupa LM. The New Visual Neurosciences. Cambridge, MA: MIT Press, 2013, with permission.)
9 to 12 Weeks of Gestation
Between 9 and 12 WG, development of intermediate and deeper choroidal vessels was observed in choroidal stroma. This development is centripetal in that it was more advanced in the posterior pole than in the equatorial choroid (Fig. 6.4). The forming vessels expressed EC markers including CD31, CD34, and CD39. Proliferation was observed in some ECs that were budding from the scleral side of the CC using CD34 and Ki67 double labeling, suggesting that intermediate vessels form by angiogenesis (9). A rather linear pattern was observed in flat CD39-labeled preparations of choroid (Fig. 6.3B) at 9 WG and then a chicken wire–like pattern of vessels with a few free CD39-positive cells between the vascular segments at 12 WG (9). This suggested that some angio-blasts were still present in the choroid (10) at 12 WG; however, the pattern of CC was approaching an adultlike lobular pattern but the density of 12 WG CC was much less than the adult CC (15).
Adult CC is fenestrated mostly on the retinal side of the CC lumens, so we investigated the presence and position of fenestrations and their components at different ages. PV-1, also called PLVAP (plasmalemmal vesicle–associated protein), is an integral membrane glycoprotein in the diaphragms of fenestrations (22,23). All vessels were negative for PV-1 at and before 12 WG. Occasional fenestrations were observed with transmission electron microscopy (TEM), but they were associated with filopodia-like structures both in and around the lumens (11). Because of their unusual position, these fenestrations were probably not functional.
We and Sellheyer and Spitznas observed that the CC at 9 WG was composed of aggregates of progenitor cells with only slit-like lumens (13). We observed some cells adventitial to cells lining the primitive lumens, but the two cell types were indistinguishable in ultrastructural appearance in periphery (11). Some plump progenitor cells that bordered on the lumens had tight junctions. Some Weibel-Palade bodies, organelles found only in vascular ECs, were present as well. Central choroid (area from disc to equator) had more definitive pericyte-like cells on the more developed vessels (11). Pericytes had a nuclear organization that appeared more distinct from the ECs on the lumens of more mature central blood vessels. Complex membranous infoldings that resembled filopodia processes extended into the slit-like lumens from the luminal cells. At the equator, some lumens were more open and the filopodia appeared to touch erythrocytes in the lumen; the plasma membranes of the two cells could not be discerned. Basal lamina was not observed around these developing vessels (11).
14 to 16 Weeks of Gestation
At 14 WG, pericyte-like cells, progenitors in the abluminal position, formed peg-and-socket–like contacts with ECs lining the lumen, a characteristic of normal adult microvasculature (11). Maturation of these abluminal cells was evaluated by localizing two pericyte markers: NG2, a glycosaminoglycan present on the surface of pericytes, and alpha smooth muscle actin (aSMA), present in mature pericytes and smooth muscle cells (SMCs). NG2 immunoreactivity was very prominent at 14 WG but there was limited aSMA immunoreactivity.
PV-1 was present at low levels in some areas of CC at 16 WG, suggesting the presence of some fenestrations. This was confirmed with TEM, which showed a few fenestrations in the CC but these were not continuous in the thin EC processes on the retinal side of the lumens (Fig. 6.5C). The CC in the posterior pole had the most fenestrations compared to peripheral regions and was most mature morphologically. In the broader lumens at 14 WG, the number of filopodia in lumens appeared greatly reduced compared to 11 WG (Fig. 6.5C). EC nuclei had less dense chromatin and were more oval and uniform in shape and BrMb organization was more advanced. The CC and intermediate choroidal vessels, which were more abundant at 14 WG, had intense APase and prominent CA IV and eNOS immunoreactivity.
21 to 22 Weeks of Gestation
Three layers of blood vessels, as demonstrated with EC markers, were apparent at 21 WG within the posterior pole (Fig. 6.4C). Short rudimentary inner segments were present at the outermost portion of the neuroblastic layer, providing the first evidence of PR maturation (11). PV-1 immunoreactivity was present in most of the CC, but it was more intense in the posterior pole than in the periphery. However, PV-1 was uniformly intense and more apparent on the retinal side of the CC lumens in the adult human eye used as a positive control (11). eNOS was prominent in EC cytoplasm and cell membrane of all choroidal blood vessels (Fig. 6.6). nNOS was mostly nuclear in both pericytes and SMCs labeled with aSMA as well as ECs, which were double labeled with von Willebrand factor (vWf) (Fig. 6.6) (19).
CC ECs were thin and fusiform ultrastructurally at 22 WG. The CC had contiguous areas of fenestrations in the narrow endothelial processes on the retinal side of the broad lumens (Fig. 6.5D). ECs had well-formed tight junctions, Weibel-Palade body numbers had increased, and a continuous basement membrane was present. Collagen and elastin was clearly deposited in BrMb under the RPE basement membrane.
aSMA+ cells were present in the CC, as well as intermediate and large choroidal blood vessels at 22 WG. Pericytes were apparent by TEM at this age (Fig. 6.5D), NG2 was very prominent, and the pericytes were located primarily on the scleral side of the CC.
Regulation of Vascular Development
Vascular development in many tissues, including the retina, is known to be controlled by vascular endothelial cell growth factor (VEGF)-A (24). The most critical isoform of VEGF-A in pathologic angiogenesis is thought to be the VEGF165 isoform (25). Recently, it has been reported that VEGF-A has two groups of splice variants from the VEGF-A gene product, VEGFxxx and VEGFxxxb, in addition to the known isoforms with varying heparinbinding affinities and molecular sizes: VEGF121’ VEGF145’ VEGF165’ VEGF189’ and VEGF206’ (26–28). The splicing of the VEGFxxxb family members, the antiangiogenic forms, is stimulated by TGF beta through splicing factor SRp55 (serine/arginine protein 55) (29). VEGFxxx, on the contrary, is proangiogenic and IGF-1 (insulin-like growth factor 1) and TNF alpha (tumor necrosis factor alpha) up-regulate splicing through ASF/SF2 (alternative splicing factor/splicing factor 2) (29,30). The two antagonistic splice variants were recently investigated in developing choroid. VEGF165 was only prominent in the forming CC during hemovasculogenesis, while VEGF165b is not present (31). VEGF165 expression increased with time in the forming vasculature and became very prominent in the basal portion of RPE and in CC at 12 WG. VEGF165 was very prominent in the basal RPE and choroidal vessels at 17 WG and VEGF165b expression was also present (31). By 21 WG, the level of the two splice variants was comparable, and both appeared to be present in the same cells (Fig. 6.7). VEGF165b localized to the nuclei of RPE and CC ECs while VEGF165 was present mostly in the cytoplasm of CC ECs and basal cytoplasm of the RPE (Fig. 6.7). Therefore, VEGF165b is only present as vascular development nears completion, while VEGF165 is prominent during hemovasculogenesis and angiogenesis.
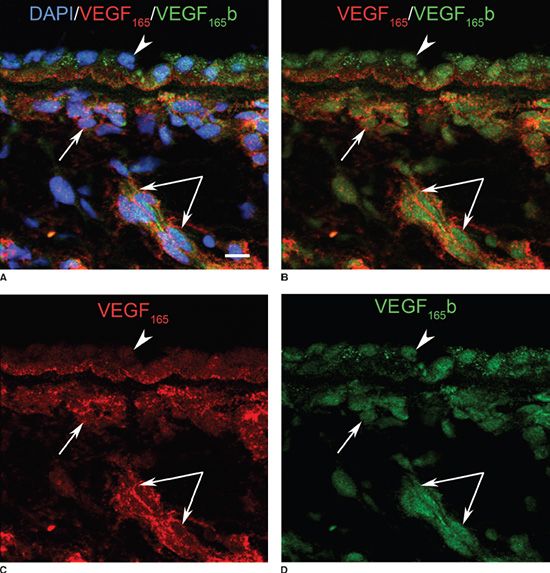
FIGURE 6.7 VEGF165 (red) (C) and VEGF165b (green) (D) coexpression (B) in the CC (arrows), deeper choroidal vessels (paired arrows), and the RPE (arrowhead). VEGF165b has an apparent nuclear localization in the endothelium of blood vessels. In the RPE, it is associated with the nuclei and in the apical portion of cells. VEGF165 is diffuse in endothelium and mostly localized to the basal portion of the RPE. Nuclei were counterstained with DAPI (blue) (D). Images are shown in single (C,D) and multiple (A,B) channels to demonstrate colabeling. (Scale bar in [A] = 10 µm.) (From Werner JS, Chalupa LM. The New Visual Neurosciences. Cambridge, MA: MIT Press, 2013, with permission.)
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


