RPE, retinal pigment epithelium.
TABLE 1.2
Time line of retinal development: organogenesis
RPE, retinal pigment epithelium.
TABLE 1.3
Time line of retinal development: differentiation

IRBP, interstitial retinol-binding protein.
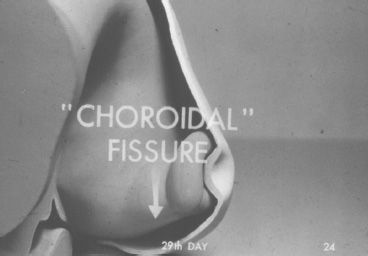
FIGURE 1.1 Four-week-old embryo. Invagination of the optic vesicle causes the formation of the optic cup and the embryonic fissure. (From Embryology of the eye [film]. San Francisco: American Academy of Ophthalmology, 1950. Courtesy of Dr. Michael Hogan.)
At the 5th week, the RPE cell is made up of two to three layers of columnar, pseudostratified pigmented cells attached one to another by junctional complexes, the anlage of the membrane of Verhoeff. The inner layer of the optic cup is the prospective sensory retina and is made of an external layer of nuclei, called the proliferative or germinative zone, and an anuclear, marginal zone. The outermost cells of the germinative zone have cilia that project toward the potential subretinal space and are joined to one another by zonulae adherens, the anlage of the external limiting membrane of the retina. The anuclear, marginal zone is covered internally by a basal lamina, the anlage of the inner limiting membrane of the retina.
The 6th week is characterized by the closure of the embryonic fissure. The closure starts at the center of the fissure and proceeds anteriorly toward the anterior rim of the optic cup and posteriorly into the optic stalk.
During the 7th week, the embryonic fissure closes completely and the anterior notch at the anterior rim of the optic cup and the posterior notch at the site of the prospective optic disc around the hyaloid artery disappear. The RPE cells extend posteriorly as the layer of the outer cells of the optic stalk, the precursor of the peripheral glial mantle of the optic nerve, which later develops into the barrier between the axons of the optic nerve and the surrounding mesenchyme (1). The RPE cells also differentiate by developing apical villi, basal infoldings, smooth and rough endoplasmic reticulum, ribosomes, premelanosomes, and melanosomes (2). In the sensory retina, germinative cells proliferate and migrate inward giving rise to the outer neuroblastic layer, the inner neuroblastic layer, and the layer of Chiewitz in between (Fig. 1.2). The outermost cells of the outer neuroblastic layer are the anlage of the photoreceptors. At the posterior pole, around the future optic disc, the innermost cells of the inner neuroblastic layer migrate inward. The first cells to migrate inward are the ganglion cells.
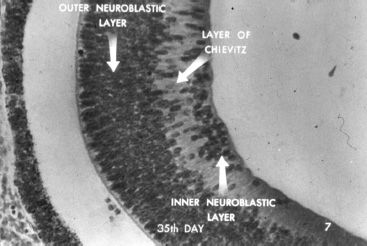
FIGURE 1.2 Seven-week-old embryo. Cells from the common neuroblastic layer of the retina arrange themselves into an outer and an inner neuroblastic layer causing the appearance of the layer of Chiewitz in between. (From Embryology of the eye [film]. San Francisco: American Academy of Ophthalmology, 1950. Courtesy of Dr. Michael Hogan.)
At the 8th week, the first ganglion cells extend axons toward the optic stalk (Fig. 1.3). These axons will develop into the future nerve fiber layer. The axons extend to and penetrate the optic stalk requiring first the apoptosis (programmed cell death) of the primitive neuroectodermal cells that fill the optic stalk (3). As more ganglion cells differentiate and axons penetrate into the optic stalk, Bergmeister papilla forms as a conic mass of glial cells covering the future optic disc area (4). Migrating retinal glioblasts, the future retinal astroglia, arrange themselves along the axons and on the internal limiting membrane (5).

FIGURE 1.3 Seven-week-old embryo. Migration inward of the ganglion cells and appearance of their axons. (From Embryology of the eye [film]. San Francisco: American Academy of Ophthalmology, 1950. Courtesy of Dr. Michael Hogan.)
At the 3rd month, the differentiation of the retina starts at the posterior pole and progresses gradually toward the periphery; the peripheral retina differentiates a short time after birth. Proliferation starts in the outer cells of the neuroblastic layer and progresses inward, while differentiation starts at the inner cells of the neuroblastic layer and extends outward. That is why the ganglion cells are the first to differentiate and the outermost cells, the anlage of the photoreceptors, are the last to differentiate (6). The differentiation in the inner neuroblastic layer manifests itself as the formation of a separate layer of ganglion cells, the appearance of glycogen granules in the cytoplasm of primitive Müller cells (7), the formation of dendrites by the ganglion cells, and the migration inwards of the amacrine cells. In the outer neuroblastic layer, it manifests itself by an inward migration of its most internal cells, the future bipolar cells, and of those external to the bipolar cells, the future horizontal cells. The migration of the future bipolar and horizontal cells causes the occlusion of the layer of Chiewitz and the formation of the external plexiform layer (the internal plexiform layer was formed as a result of the migration of the ganglion cells). The outermost cells of the outer neuroblastic layer are connected one to the other with adherent junctions, the future external limiting membrane.
At the 4th month, all the major constituents of the retina are present (Fig. 1.4). The ora serrata appears as a line of demarcation at the peripheral retina. At the posterior pole, the outer nuclear layer becomes the precursor of the rod-free zone, the future fovea. The inner plexiform layer acquires ribbon and conventional synapses. As neurons mature, so do primitive Müller cells, which acquire more glycogen, intermediate filaments, myelin-associated protein, and hyaluronic acid. The differentiation of the photoreceptors manifests by the production of F-actin and alpha-tubulin, components of the microtubules. At this point, there is not yet a subretinal space present. The most important event occurring at this stage is the appearance of retinal vessels. Cells originating inside the optic disc from the walls of the two venous channels located on either side of the hyaloid artery and from the adventitia of the hyaloid artery proliferate and migrate into the inner retina. They differentiate into endothelial cells, which first make cords and then canalize to form capillaries. These cells possess a vascular precursor marker, such as CD39, and have been named angioblasts. The initial process of retinal vascularization is attributed to the formation of de novo vessels from precursor vascular cells and is thus believed due to vasculogenesis.
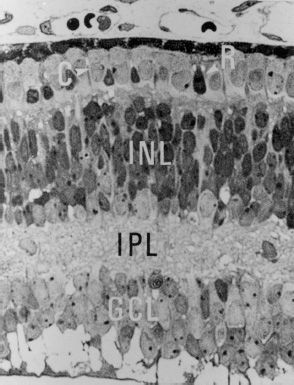
FIGURE 1.4 Four-month-old human fetus. All the major constituents of the retina are present. Cones (C), rods (R), external plexiform layer, internal nuclear layer (INL), internal plexiform layer (IPL), ganglion cell layer (GCL), nerve fiber layer, and internal limiting membrane can be clearly distinguished (×540). (Reproduced from Hollenberg MJ, Spira AW. Early development of the human retina. Can J Ophthalmol 1972;7:472–491, with permission.)
The 5th month is characterized by the conspicuous differentiation of the photoreceptors: Membrane infoldings become the tubular structures of the outer segments and rhodopsin and S antigen appear (8). The differentiation of the apical surfaces of the photoreceptors and of the apical villi of the RPE cells leads to breakdown of junctions between these cells and the formation of the subretinal space (Fig. 1.5). Photoreceptors secrete interstitial retinol-binding protein (IRBP) into the subretinal space. After the photoreceptors develop, horizontal cells become conspicuous in a row. At the internal layers of the retina, amacrine and ganglion cells are apparent in their definitive locations. Apoptosis of cells, including ganglion cells, causes the appearance of cell debris that is phagocytosed by surrounding cells, not believed to be macrophages, as retinal vascularization (the believed source of macrophages) has not yet completely developed. The cell loss is not uniform, being more pronounced at the periphery and causing a centrifugal gradient in the distribution of ganglion cells in the fetal retina. The pars plana begins to develop and renders the ora serrata more clearly distinguishable. Retinal vascularization progresses rapidly. Newly formed capillaries extend peripherally and form arteries and veins; the arteries and veins present at the optic disc become the central retinal artery and veins. In capillaries, cells in contact with the blood flow become endothelial cells and those surrounding endothelial cells become pericytes. Processes of astrocytes attach to the collagenous matrix surrounding the capillaries (see also Chapter 6).
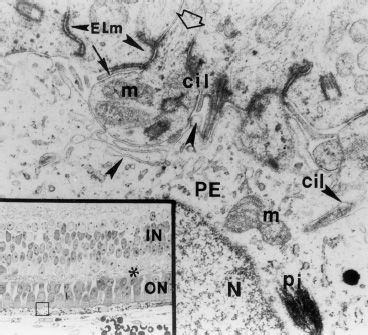
FIGURE 1.5 Five-month-old human fetus. The inset shows all the layers of the retina. Amacrine cells with large, light-colored nuclei; Müller cells with dark, angular nuclei; bipolar cells with small, oval nuclei of medium density; horizontal cells with large nuclei. ON, outer nuclear layer; IN, inner nuclear layer; asterisk, outer plexiform layer. The main figure shows the pigment epithelium–photoreceptor contact area. The blank arrow points to the beginning of an inner segment. The thin solid arrow indicates villi of a retinal pigment epithelial (PE) cell. There are junctions between the RPE cells and the photoreceptors. The subretinal space is not yet present. cil, cilium; ELm, external limiting membrane; m, mitochondria; N, nucleus of pigment epithelial cell; pi, pigment granule; arrow heads, microvilli of pigment epithelial cell (×24,000). (Reproduced from Ozanics J. Prenatal development of the eye and its ednexae. In: Ocular anatomy, embryology, and teratology. Philadelphia, PA: Harper & Row, 1982:11–96, with permission.)
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


