As mammals evolved, the larynx accrued added importance in new activities such as effectuating intra-abdominal pressure control during the transition from egg-laying to birthing and control of intrathoracic stabilization as movement of the upper limb required rib stabilization during climbing. The proper development of the larynx during the embryonic period is crucial to its later normal function. The permanent intersection of the respiratory and digestive pathways has created a de novo aerodigestive tract; a first of its kind in mammals. The lowered position of the human larynx has provided a greatly expanded supralaryngeal portion of the pharynx.
Key points
- •
As mammals morphed from their amphibian and reptilian ancestors, their larynx accrued added importance in new activities such as effectuating intra-abdominal pressure control during the transition from egg-laying to birthing and control of intrathoracic stabilization as movement of the upper limb required rib stabilization during climbing.
- •
The proper development of the larynx during the embryonic period is crucial to its later normal function, with miscues during this phase of development resulting in a range of serious congenital, often life-incompatible, anomalies.
- •
The permanent intersection of the respiratory and digestive pathways has created a de novo “aerodigestive” tract, a first of its kind in mammals.
- •
The lowered position of the human larynx has provided one major positive aspect: a greatly expanded supralaryngeal portion of the pharynx that enables speech production.
- •
The panoply of swallowing problems seen today was starting to come to the fore as the unique human ADT evolved.
Of all human activities, the function we probably least appreciate is swallowing. We must breathe and are always acutely aware of our breathing patterns. The slightest disruption or sniffle causes immediate reactions. Bathroom needs and activities are also high on our list (particularly as we age; can you say “prostate”). And we protect and monitor our senses – from smell to sight to hearing and balance – scrupulously. We infrequently, however, think about the many manifestations of swallowing, or of a swallow, unless, of course, one cannot do it.
The ability to swallow is arguably the “Rodney Dangerfield” of human aerodigestive functions: it receives little respect. The movement of a bolus from initial contact with our external lips through the various oral and pharyngeal portals, sphincters, and way-stations en route to our gastro-esophageal processing chambers is given nary a thought. Whether we swallow saliva or transport a heavy density bolus of meat, whether we are young or old, male or female, from New York or New Delhi, the expectation is that our swallowing mechanism will function, and function constantly and well.
We humans arguably sit at the pinnacle of the mammalian world. Linnaeus unabashedly anointed our biological order, Primates, the “firsts.” Our feline or cetacean brethren may take some umbrage, but since they can’t speak we will never know. We are the masters of our planet and our evolution has endowed us with incomparable cognitive and communicative abilities that have allowed us the pre-eminence of culture and mastery of our world.
As our kind meandered through the corridors of evolution our basic body plan morphed and changed through genetics and environmental necessity to arrive at the masterful visage reflected in the mirror each morning. We evolved our distinctive habitually bipedal mode of locomotion, derived dentognathic arrangements commensurate with our dietary palate and unrivaled intricacies of our brain in the entangled evolutionary dance. Tied with all this, was an equally remarkable re-arrangement and functional redesign of the region of our interests, the aerodigestive tract, the unwieldy handle most have come to use for this mixed-use realm.
Indeed, we humans are arguably the only mammal – assuredly, only living primate (prosimians, monkeys, apes and us) – that has a constantly communal respiratory, digestive, vocal, aeration, olfactory and pressure control “tract.” The very fact that we have no scientifically precise name for this noisy confluence of functions – it is neither purely respiratory, purely digestive, nor purely vocal – speaks to how the region has morphed and changed in starts and stops during eons of our evolution. All these changes have produced the unique substrate that allows for our distinctive modes of swallowing.
The aerodigestive tract among mammals
One cannot understand the function of the human “aerodigestive” tract (ADT), let alone its evolution or unique ontogenetic development, without first viewing it in a comparative mammalian context. To gain some handle on the ADT it is necessary to have a focal point for that discussion, and that touchstone is the larynx – arguably one of the key structures in mammalian anatomy and function. As air-breathing mammals, the need to protect the lungs from incursions became paramount. Although we have ascended the evolutionary ladder, the primary function of our larynx remains true to its heritage: it is still essentially a valve, regulating and guarding the airway. As mammals morphed from their amphibian and reptilian ancestors their larynx also accrued added importance in new activities such as effectuating intra-abdominal pressure control during the transition from egg-laying to birthing, and control of intrathoracic stabilization as movement of the upper limb required rib stabilization during climbing. Although the larynx of diverse mammalian species share many homologous components, the specifics of structure for larynges of species that inhabit often vastly differing environments have been modified extensively during the course of evolution.
While mammals exhibit great variation in body plan, the general template for their throat region is remarkably similar ( Fig. 1 ). A hallmark of most mammals is that the epiglottis can make contact, or overlap the soft palate, during both normal respiration and deglutition. This is effectuated in most species due to a larynx positioned, at all stages of postnatal development, relatively “high” in the neck when related to the basicranium and/or cervical vertebrae. Its position, measured from the cranial aspect of the epiglottis to the caudal border of the cricoid cartilage, corresponds to the level of the basiocciput or first cervical vertebra (C1) to the third or fourth cervical vertebrae (C3 or C4) in most terrestrial mammals. The hyoid bone and associated suprahyoid and infrahyoid muscles (i.e., muscles largely responsible for raising or lowering the larynx) are also relatively high. The tongue at rest lies almost entirely within the oral cavity, with no portion of it forming part of the anterior pharyngeal wall. Because of this high position of the larynx, the supralaryngeal region of the pharynx is noticeably small; the pharynx has little or no oral portion and significantly reduced nasal and laryngeal segments. Inferiorly, the striated muscle fibers of the pharynx blend with the longitudinal striated fibers of the esophagus to form a continuous functional unit.
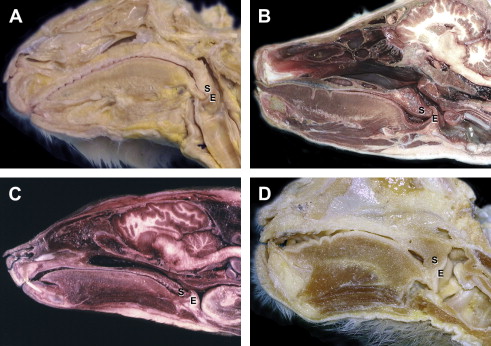
As noted, the high position of the larynx enables the epiglottis to pass upward behind the soft palate and lock the larynx directly into the nasopharynx. This configuration provides a direct air channel from the external nares through the nasal cavities, nasopharynx, larynx, and trachea to the lungs. Liquids, and in some species even chewed or solid material, can pass on either side of the interlocked larynx and nasopharynx by way of the isthmus faucium, through the piriform sinuses to the esophagus, following the so-called “lateral food channels.” This anatomic configuration allows streams of liquid or semisolid food to be transmitted around each side of the larynx during swallowing, while also maintaining patency of the laryngeal airway. Two largely separate pathways are created: a digestive tract from the oral cavity to the esophagus and a respiratory tract from the nose to the lungs. This arrangement confers on mammals the ability to use these two pathways simultaneously, including enabling simultaneous breathing while: (1) young are suckling, (2) ruminants are regurgitating cud, (3) carnivores have their mouth clamped tightly closed around the neck of their prey, and (4) a variety of animals use the mouth as a tool (e.g., beavers gnawing trees, felines or rodents grasping and carrying young). In addition, as many mammals are macrosmatic (i.e., largely dependent on olfaction for communication with their environment), the two-tube system is particularly valuable as this arrangement allows, for example, grazing or drinking herbivores to simultaneously detect the scent of a predator.
Although the larynx is consistently high in most mammals, its exact position and the extent of its placement in the nasopharynx can vary considerably among species. For example, studies of cetaceans (i.e., whales, dolphins, porpoises) have shown that the larynx is positioned so high (rostral) that it is no longer solely in the neck but rather lies largely within the head. The epiglottis lies under the presphenoidal synchondrosis, while the caudal border of the cricoid is positioned approximately opposite the occipital condyles or perhaps C1. The compressed cervical vertebrae characteristic of these mammals makes it difficult to determine a corresponding cervical position, as there is very little neck region for the larynx to occupy. The anterior portion of the larynx is elongated into a tube that is encircled by a strong palatopharyngeal sphincter muscle (homologous to the soft palate and palatopharyngeal arch in humans). This sphincter grips the laryngeal tube and keeps its aditus intranarial, thus effectively sealing the respiratory tract from the digestive route. Although they exhibit the baseline mammalian pattern of a high larynx, odontocetes appear to have exaggerated it by placing the larynx even higher (or more rostral) than their terrestrial relatives. This extra high position ensures these mammals of a larynx fitted snugly into the nasopharynx, and indeed may make it permanently intranarial—that is, it is not normally retracted from its position behind the soft palate. This arrangement may allow them to swallow whole fish while keeping the airway patent for generating sounds for communication or navigation (echolocation).
Although cetaceans demonstrate an example of larynges that have both migrated cranially and elongated their rostral cartilages, some terrestrial species have larynges that have expanded their caudal components (e.g., thyroid cartilage) so that they appear to extend considerably into the neck. For example, some male artiodactyls (red and fallow deer, Mongolian gazelle) exhibit particularly large larynges that seem to be located more caudally in the neck compared with other related species. However, although the larynges of these animals are elongated, they still retain roughly the same position opposite the cervical vertebrae as most other terrestrial mammals (extending from the basiocciput to C2-C3). Maintenance of this typical mammalian position is due to concomitant elongation of the cervical vertebrae. These animals also exhibit an elongated and elastic velum (red and fallow deer) and an elongated epiglottis (Mongolian gazelle) that appear to assist in epiglottic/palatal contact and, therefore, the maintenance of the two-pathway system. Thus, while larynges differ considerably in position, the ancestral “two-tube” configuration is essentially maintained. These animals have modified a basic plan; they have not changed it.
Postmortem dissections and a range of imaging studies, including cineradiography, CT, and MR, of our closest relatives the nonhuman primates show that their upper respiratory anatomy is also similar to the general mammalian pattern. As in other mammals, nonhuman primates exhibit a larynx positioned high in the neck, usually corresponding to the first to third cervical vertebrae ( Fig. 1 ). This position allows for epiglottic-soft palate apposition and the possibility of an intranarial larynx, thus providing a continuous airway from the nose to the lungs, while the alimentary tract passes around the larynx en route to the esophagus. Cineradiographic studies have confirmed that nonhuman primates exhibit mostly separate respiratory and digestive routes and the ability to breathe and swallow almost simultaneously. Because of this configuration, nonhuman primates, like other mammals, appear strongly, if not totally, dependent on nasal breathing. As occurs in many mammals, the connection between the epiglottis and the soft palate can be broken, as the larynx exhibits extensive mobility and can be transiently lowered. This can occur for a number of reasons, including some vocalizations, swallowing certain foods (e.g., a large bolus of meat), or due to disease.
Although this anatomic arrangement may enable almost simultaneous breathing and swallowing, it severely limits the array of sounds an animal can produce. The high position of the larynx means that only a small supralaryngeal portion of the pharynx exists. In turn, only a very reduced area is available to modify the initial sounds generated at the vocal folds. Due to this limitation, most mammals therefore depend primarily on altering the shape of the oral cavity and lips to modify laryngeal sounds. Although some animals can approximate some human speech sounds, they are anatomically incapable of producing the range of sounds necessary for human speech.
The aerodigestive tract among mammals
One cannot understand the function of the human “aerodigestive” tract (ADT), let alone its evolution or unique ontogenetic development, without first viewing it in a comparative mammalian context. To gain some handle on the ADT it is necessary to have a focal point for that discussion, and that touchstone is the larynx – arguably one of the key structures in mammalian anatomy and function. As air-breathing mammals, the need to protect the lungs from incursions became paramount. Although we have ascended the evolutionary ladder, the primary function of our larynx remains true to its heritage: it is still essentially a valve, regulating and guarding the airway. As mammals morphed from their amphibian and reptilian ancestors their larynx also accrued added importance in new activities such as effectuating intra-abdominal pressure control during the transition from egg-laying to birthing, and control of intrathoracic stabilization as movement of the upper limb required rib stabilization during climbing. Although the larynx of diverse mammalian species share many homologous components, the specifics of structure for larynges of species that inhabit often vastly differing environments have been modified extensively during the course of evolution.
While mammals exhibit great variation in body plan, the general template for their throat region is remarkably similar ( Fig. 1 ). A hallmark of most mammals is that the epiglottis can make contact, or overlap the soft palate, during both normal respiration and deglutition. This is effectuated in most species due to a larynx positioned, at all stages of postnatal development, relatively “high” in the neck when related to the basicranium and/or cervical vertebrae. Its position, measured from the cranial aspect of the epiglottis to the caudal border of the cricoid cartilage, corresponds to the level of the basiocciput or first cervical vertebra (C1) to the third or fourth cervical vertebrae (C3 or C4) in most terrestrial mammals. The hyoid bone and associated suprahyoid and infrahyoid muscles (i.e., muscles largely responsible for raising or lowering the larynx) are also relatively high. The tongue at rest lies almost entirely within the oral cavity, with no portion of it forming part of the anterior pharyngeal wall. Because of this high position of the larynx, the supralaryngeal region of the pharynx is noticeably small; the pharynx has little or no oral portion and significantly reduced nasal and laryngeal segments. Inferiorly, the striated muscle fibers of the pharynx blend with the longitudinal striated fibers of the esophagus to form a continuous functional unit.
As noted, the high position of the larynx enables the epiglottis to pass upward behind the soft palate and lock the larynx directly into the nasopharynx. This configuration provides a direct air channel from the external nares through the nasal cavities, nasopharynx, larynx, and trachea to the lungs. Liquids, and in some species even chewed or solid material, can pass on either side of the interlocked larynx and nasopharynx by way of the isthmus faucium, through the piriform sinuses to the esophagus, following the so-called “lateral food channels.” This anatomic configuration allows streams of liquid or semisolid food to be transmitted around each side of the larynx during swallowing, while also maintaining patency of the laryngeal airway. Two largely separate pathways are created: a digestive tract from the oral cavity to the esophagus and a respiratory tract from the nose to the lungs. This arrangement confers on mammals the ability to use these two pathways simultaneously, including enabling simultaneous breathing while: (1) young are suckling, (2) ruminants are regurgitating cud, (3) carnivores have their mouth clamped tightly closed around the neck of their prey, and (4) a variety of animals use the mouth as a tool (e.g., beavers gnawing trees, felines or rodents grasping and carrying young). In addition, as many mammals are macrosmatic (i.e., largely dependent on olfaction for communication with their environment), the two-tube system is particularly valuable as this arrangement allows, for example, grazing or drinking herbivores to simultaneously detect the scent of a predator.
Although the larynx is consistently high in most mammals, its exact position and the extent of its placement in the nasopharynx can vary considerably among species. For example, studies of cetaceans (i.e., whales, dolphins, porpoises) have shown that the larynx is positioned so high (rostral) that it is no longer solely in the neck but rather lies largely within the head. The epiglottis lies under the presphenoidal synchondrosis, while the caudal border of the cricoid is positioned approximately opposite the occipital condyles or perhaps C1. The compressed cervical vertebrae characteristic of these mammals makes it difficult to determine a corresponding cervical position, as there is very little neck region for the larynx to occupy. The anterior portion of the larynx is elongated into a tube that is encircled by a strong palatopharyngeal sphincter muscle (homologous to the soft palate and palatopharyngeal arch in humans). This sphincter grips the laryngeal tube and keeps its aditus intranarial, thus effectively sealing the respiratory tract from the digestive route. Although they exhibit the baseline mammalian pattern of a high larynx, odontocetes appear to have exaggerated it by placing the larynx even higher (or more rostral) than their terrestrial relatives. This extra high position ensures these mammals of a larynx fitted snugly into the nasopharynx, and indeed may make it permanently intranarial—that is, it is not normally retracted from its position behind the soft palate. This arrangement may allow them to swallow whole fish while keeping the airway patent for generating sounds for communication or navigation (echolocation).
Although cetaceans demonstrate an example of larynges that have both migrated cranially and elongated their rostral cartilages, some terrestrial species have larynges that have expanded their caudal components (e.g., thyroid cartilage) so that they appear to extend considerably into the neck. For example, some male artiodactyls (red and fallow deer, Mongolian gazelle) exhibit particularly large larynges that seem to be located more caudally in the neck compared with other related species. However, although the larynges of these animals are elongated, they still retain roughly the same position opposite the cervical vertebrae as most other terrestrial mammals (extending from the basiocciput to C2-C3). Maintenance of this typical mammalian position is due to concomitant elongation of the cervical vertebrae. These animals also exhibit an elongated and elastic velum (red and fallow deer) and an elongated epiglottis (Mongolian gazelle) that appear to assist in epiglottic/palatal contact and, therefore, the maintenance of the two-pathway system. Thus, while larynges differ considerably in position, the ancestral “two-tube” configuration is essentially maintained. These animals have modified a basic plan; they have not changed it.
Postmortem dissections and a range of imaging studies, including cineradiography, CT, and MR, of our closest relatives the nonhuman primates show that their upper respiratory anatomy is also similar to the general mammalian pattern. As in other mammals, nonhuman primates exhibit a larynx positioned high in the neck, usually corresponding to the first to third cervical vertebrae ( Fig. 1 ). This position allows for epiglottic-soft palate apposition and the possibility of an intranarial larynx, thus providing a continuous airway from the nose to the lungs, while the alimentary tract passes around the larynx en route to the esophagus. Cineradiographic studies have confirmed that nonhuman primates exhibit mostly separate respiratory and digestive routes and the ability to breathe and swallow almost simultaneously. Because of this configuration, nonhuman primates, like other mammals, appear strongly, if not totally, dependent on nasal breathing. As occurs in many mammals, the connection between the epiglottis and the soft palate can be broken, as the larynx exhibits extensive mobility and can be transiently lowered. This can occur for a number of reasons, including some vocalizations, swallowing certain foods (e.g., a large bolus of meat), or due to disease.
Although this anatomic arrangement may enable almost simultaneous breathing and swallowing, it severely limits the array of sounds an animal can produce. The high position of the larynx means that only a small supralaryngeal portion of the pharynx exists. In turn, only a very reduced area is available to modify the initial sounds generated at the vocal folds. Due to this limitation, most mammals therefore depend primarily on altering the shape of the oral cavity and lips to modify laryngeal sounds. Although some animals can approximate some human speech sounds, they are anatomically incapable of producing the range of sounds necessary for human speech.
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


