PRINCIPAL CAUSES
There are four principal groups of bacteria that are most frequently responsible (
3): Micrococcaceae (
Staphylococcus, Micrococcus), the
Streptococcus species, the
Pseudomonas species, and the Enterobacteriaceae (
Citrobacter, Klebsiella, Enterobacter, Serratia, Proteus). However, virtually any bacteria can potentially cause keratitis under certain favorable conditions. A classification of the medically important bacteria based on type, Gram stain, and oxygen requirements for growth is provided in
Table 13.1-1. The flora of the ocular surface and the environment in which the person lives influence the type of infection that develops. Ambient temperature and humidity have a major role in determining the microorganisms found in the environment. Pathogens isolated from cases of microbial keratitis vary among geographic locations according to the local climate and occupational risk factors. Differences were reported in isolates from patients with suppurative keratitis from Ghana and southern India, both of which are at similar tropical latitudes (
4). There were differences found in the bacterial isolates, with
Pseudomonas species the most frequent isolate from Ghana and
Streptococcus species the most common isolate from southern India. Geographic variations also exist in the relative frequency of different bacterial organisms as causative agents in keratitis in the United States. Pneumococcus (
Streptococcus pneumoniae) was a frequently encountered causative organism of bacterial keratitis in previous clinical reports because of its association with chronic dacryocystitis (
5). Pneumococcus has decreased in frequency as a causative organism in developed countries with available effective antibiotics and with refinements in techniques for dacryocystorhinostomy. In developing countries, the pneumococcus may remain an important cause of infectious corneal ulceration (
6,
7,
8). Other gram-positive organisms, especially among the
Staphylococcus species, continue to be the most commonly isolated causes of bacterial keratitis.
Staphylococcus aureus is among the most frequent causative organisms in bacterial keratitis in the Northern and North Eastern United States and Canada, both in normal hosts and in compromised corneas (
9). In Great Britain, the most common organisms isolated in bacterial keratitis are
S. aureus, S. pneumoniae, Pseudomonas, and
Moraxella (
10).
Once an infrequent isolate,
Pseudomonas species became increasingly isolated as causative organisms in keratitis.
Pseudomonas has been reported as the most commonly
isolated organism in the southern regions of the United States (
11,
12). The increasing prevalence of
Pseudomonas in otherwise healthy individuals was found largely in association with daily- or extended-wear soft contact lenses (
13,
14,
15,
16,
17,
18,
19).
Pseudomonas is widely distributed in nature and is capable of being a contaminant in the hospital environment, in fluorescein sodium solutions, in cosmetics, and organic carbon substances (
20).
Pseudomonas has long been recognized as a potential opportunistic pathogen in burn patients, patients with altered consciousness, patients with exposure, and in individuals maintained on mechanical ventilatory assistance.
Shifting trends in bacterial keratitis in south Florida were observed and reported during the 1990s (
21).
S. aureus and
Pseudomonas aeruginosa represented 19.4% and 25.7%, respectively, of the total bacterial isolates during this period. However, a gradual increase was documented in the number of
S. aureus keratitis isolates (29% of grampositive organisms in 1990 versus 48% in 1998,
p = .01) coupled with a decrease in the number of
P. aeruginosa isolates (54% of gram-negative organisms in 1990 vs. 46% in 1998). A decrease in the incidence of contact lens-associated keratitis and
P. aeruginosa isolates in this group of patients was documented.
Serratia marcescens and
P. aeruginosa were most commonly isolated in contact lens-associated keratitis (18% each).
Distribution and shifting trends of bacterial keratitis in north China were reported over the period of 1989 to 1998 (
22). The data of 2220 corneal isolates were reviewed retrospectively. Positive culture was recovered in 490 isolates. Gram-positive cocci and gram-negative bacilli represented 51% and 39.4%, respectively.
P. aeruginosa was the most common pathogen (32.2%). A gradual increase in the percentage of gram-positive cocci was coupled with a decrease of gram-negative bacilli.
P. aeruginosa and coagulase-negative
Staphylococcus were the most common pathogens in bacterial keratitis in north China.
Moraxella, a gram-negative aerobic bacterium, may be a causative agent of keratitis in malnourished individuals and patients with alcoholism, diabetes, and other debilitating conditions (
5,
9,
20).
Moraxella may also cause bacterial keratitis in healthy patients.
Moraxella is present in 0.3% of normal eyelids and in up to 0.8% of inflamed eyelids (
23).
The indigenous bacteria, including coagulase-negative
Staphylococcus, Corynebacterium, and
Propionibacterium species, are increasing in frequency of isolation from bacterial keratitis cases because of their proximity to the cornea and the potential for direct inoculation (
24). There are a number of species of coagulase-negative
Staphylococcus. Although
Staphylococcus epidermidis has been reported as the most frequently isolated from ocular sources, with newer, highly specific laboratory identification systems, it is possible that other coagulase-negative staphylococci may be increasingly implicated as causative organisms. In a multicenter, prospective, randomized comparative study of therapy for bacterial keratitis, coagulase-negative
Staphylococcus species were among the most frequently encountered causative organisms (
25).
Since the development of a vaccine against
Corynebacterium diphtheriae, isolation of the organism from the eye is rare. Corneal involvement with
C. diphtheriae is infrequent and typically observed after a reaction to diphtheria toxin (
26). By virtue of the activity of its toxin,
C. diphtheriae may penetrate intact corneal epithelium to produce keratitis.
Other
Corynebacterium species have been recovered from infectious corneal ulcerations. The precise causative role of these
Corynebacterium species has occasionally been uncertain. Cases in which
Corynebacterium species have been the sole organisms isolated have increased the awareness of their potential as important causative organisms of keratitis (
27,
28).
Corynebacterium xerosis and
Corynebacterium striatus are the species that have been most commonly recovered.
C. xerosis may be a coinfectious agent frequently associated with
Moraxella infections.
Propionibacterium species are non-spore-forming anaerobic bacteria that have been implicated after sequestration in the capsular bag as a specific cause of chronic pseudophakic endophthalmitis (
29).
Propionibacterium species are also capable of causing keratitis in healthy and compromised corneas.
Gram-positive aerobic bacilli are widespread in nature and are usually of low virulence. They may produce infections of the cornea when host resistance is lowered (
30).
Bacillus cereus has been associated with severe posttraumatic endophthalmitis after penetrating injury with vegetable or metallic matter. Specific enzymes and toxins, including hemolysins, enterotoxins, and emetic toxin, are produced by this organism. These virulence factors may be particularly destructive to the retina and responsible for fulminant destruction (
31,
32).
B. cereus keratitis may follow foreign body injury (
33). Other
Bacillus species that have been implicated as causative agents of bacterial keratitis include
Bacillus brevis, Bacillus coagulans, Bacillus laterosporus, Bacillus licheniformis, and
Bacillus thuringiensis (
30,
34,
35).
Organisms less frequently associated with keratitis include
Serratia (
36),
Proteus, and
Azotobacter (
37).
S. marcescens corneal ulcerations have been associated with contact lens wear (
36) and contaminated eyedrops (
38).
Other aerobic gram-negative organisms capable of causing keratitis include Neisseria gonorrhoeae and Neisseria meningitidis. N. gonorrhoeae may produce consecutive keratitis during an episode of untreated or inadequately treated purulent conjunctivitis. Neisseria species can cause marked conjunctival chemosis and infiltration with polymorphonuclear neutrophils. The edematous infiltrated conjunctiva may then drape over the peripheral cornea, resulting in exposure to neutrophilic lytic enzymes capable of producing epithelial ulceration. The pace of inflammation may result in rapid stromal thinning and perforation. It is imperative to verify the presence of the gonococcus by culture because Acinetobacter, which is morphologically identical on smears to the gonococcus, can also cause corneal perforation.
Non-spore-forming anaerobic bacteria are principal flora of the skin, oral cavity, gastrointestinal tract, and other mucous membranes. These anaerobic bacteria are capable of causing keratitis and other ocular infections (
39). The most frequently isolated organisms are
Peptostreptococcus, Peptococcus, and
Propionibacterium (
40,
41). Spore-forming anaerobic bacteria such as
Clostridium species may be rarely encountered as causes of infectious corneal ulceration, especially after contamination with soil (
42). The presence of
Clostridium infection may be suggested by gas bubbles observed in the corneal tissue or anterior chamber (
43).
Listeria monocytogenes is a facultatively anaerobic, gram-positive rod that is infrequently isolated as a cause of severe, suppurative keratitis, especially among animal caretakers (
44). The organism is an intracellular pathogen which may also cause endophthalmitis with pigment dispersion, dark hypopyon, and elevated intraocular pressure (
45).
In studies that used adequate methods for recovery of anaerobes, these bacteria were isolated from approximately a third of patients with conjunctivitis, half of the time in pure culture (
46). The predominant recovered anaerobes were
Clostridium species, gram-negative anaerobic bacilli, and
Peptostreptococcus species. Anaerobic bacteria were also recovered from patients who wore contact lenses and had conjunctivitis. Anaerobic bacteria were also reported in cases of keratitis (
46). The most frequently recovered anaerobes were
Propionibacterium, Peptostreptococcus, Clostridium, Prevotella, and
Fusobacterium species. The most frequently recovered anaerobes from dacryocystitis were
Peptostreptococcus, Propionibacterium, Prevotella, and
Fusobacterium species.
Members of the actinomycetes and related organisms, including
Nocardia, are known to resemble bacteria more than fungi. These gram-positive, branching filamentous organisms have been causally linked with infections of the lacrimal system, yet are rarely encountered as isolates from bacterial keratitis (
47).
Nocardia organisms are ubiquitously present in soil and may cause infectious corneal ulceration after trauma (
48).
Primary tuberculous keratitis is now extremely rare. Nontuberculous mycobacteria, including
Mycobacterium fortuitum (
49,
50),
Mycobacterium chelonae (
51,
52),
Mycobacterium gordonae (
53), and
Mycobacterium avium-intracellulare (
54), are all capable of causing an indolent, intractable keratitis, especially after injury with a foreign body or after office surgical procedures (
55). A case of keratitis with extension into the sclera has been reported due to
Mycobacterium marinum (
56).
Acute and delayed-onset mycobacterial keratitis after laser
in situ keratomileusis (LASIK) has been reported with increasing frequency and in clusters or outbreaks at some outpatient laser vision correction centers around the world (
57). Infectious keratitis after LASIK is a potentially vision-threatening complication (
58). Onset of symptoms varies depending on causative agents. Furthermore, atypical organisms in the interface or beneath the flap can pose both diagnostic and therapeutic dilemmas. Location in the interface can make it more difficult to culture the organisms and prevent adequate penetration of topical antibiotics. In seven patients,
Mycobacterium szulgai keratitis developed from 7 to 24 weeks after LASIK (
59). Nontuberculous mycobacteria were identified 1 month after the flaps were cultured. Pulsed-field gel electrophoresis (PFGE) was used to type the
isolates, and treatment was modified based on susceptibilities.
M. szulgai was identified in five patients for whom cultures were performed, but response to empirical therapy based on cultures proved unsatisfactory. The keratitis resolved in all patients with treatment including clarithromycin based on susceptibilities. Medical therapy was sufficient, although one patient required flap amputation. Six of seven patients recovered best corrected visual acuity (BCVA), whereas one patient lost one line of BCVA. Two patients lost one line of postoperative uncorrected visual acuity (UCVA), two patients gained one line of UCVA, and three patients recovered postoperative UCVA. PFGE analysis revealed that the
M. szulgai strains were identical, and the infection source was contaminated ice used to chill syringes for saline lavage. Nontuberculous mycobacterial keratitis after LASIK is a diagnostic and management challenge. This cluster and other outbreaks underscore the importance of adherence to sterile protocol during LASIK.
The incidence of ocular lesions of leprosy varies from approximately 15% of patients with tuberculoid leprosy to approximately 100% of the patients with long-standing lepromatous leprosy (
60).
Mycobacterium leprae is the causative agent of keratitis and may invade along peripheral corneal nerves (
61,
62).
Spirochetal infection with
Treponema pallidum may result in a nonsuppurative stromal keratitis as a rare complication of acquired primary or secondary syphilis (
63). Nonsyphilitic spirochetal infection of the cornea may occur in Lyme disease. A nonsuppurative, deep stromal keratitis similar to that observed in syphilis may be caused by the spirochete
Borrelia burgdorferi (
64). Organisms less frequently causing bacterial keratitis include
Aeromonas hydrophilia (
65),
Moraxella catarrhalis (
66),
Pasteurella multocida (
67), and complex, gram-negative bacilli, including
Methylobacterium (Pseudomonas) mesophilica, Capnocytophaga species, and
Alcaligenes xylosoxidans subsp.
denitrificans.
RISK FACTORS
Bacterial keratitis typically occurs in eyes having one or more predisposing risk factors (
68). Although the eye is constantly exposed to a large number of bacteria, it is a tribute to the natural host defense mechanisms that only a small portion of these microorganisms result in corneal infection. The microflora of the eye is diverse and continuously changing (
69). To maintain ocular health, the various pathogenic bacteria on the ocular surface must be eliminated before proliferation and infection. There are several mechanisms to protect the surface of the eye from infectious agents. The eyelids provide a physical barrier to protect against organisms gaining direct access to the eye. The preocular tear film provides natural irrigation to remove organisms (
70). The tear film contains secretory immunoglobulins, complement components, and various enzymes, including lysozyme, lactoferrin (
71), betalysins, orosomucoid, and ceruloplasmin (
72). The mucin layer of the tear film produced by goblet cells acts as a mechanical protective barrier that can trap and remove potentially pathogenic organisms. The normal ocular flora provide a balance to help prevent overgrowth of exogenous organisms. The conjunctiva contains subepithelial mucosal-associated lymphoid tissue with a collection of lymphoid cells having specific defensive functions (
73). T-cell lymphocytes are present in the conjunctiva in a ratio to B lymphocytes of approximately 20:1 in the substantia propria (
74). Polymorphonuclear neutrophils (PMN) may be attracted to the cornea by chemotactic factors. Immunoglobulin G (IgG) in tears may assist by binding to bacteria, fixing complement, or enhancing phagocytosis. Cell-mediated immunity has an important role in the ocular defense against certain bacteria, especially intracellular pathogens. The cellular immune response may be initiated on the ocular surface by a specialized type of dendritic cell known as a
Langerhans cell. The Langerhans cells are found in high density at the sclerocorneal limbus and diffusely throughout the bulbar conjunctiva (
75). There are normally few Langerhans cells present in the central cornea (
76). Langerhans cell surface expresses human leukocyte antigen DR (class II histocompatibility antigen), Ia antigen, and receptors for complement and the Fc portion of immunoglobulin (
75). The Langerhans cells have an important role in the processing of bacterial antigens and their presentation to T cells.
Perhaps the most important defense barrier for the cornea is an intact epithelial layer. Most corneal infections result from trauma to the corneal epithelium. Alteration of any of the local or systemic defense mechanisms may also predispose the host to corneal infection. Eyelid abnormalities, including ectropion with exposure, entropion with trichiasis, or lagophthalmos, may be local factors contributing to corneal infection. Abnormalities of the preocular tear film, including aqueous tear layer insufficiency, mucin layer deficiencies from goblet cell loss or dysfunction, and lipid layer instability, may predispose to bacterial keratitis. Lacrimal drainage obstruction may interfere with the lubricating mechanical defense function. The inappropriate use of topical antibiotics could eliminate the natural protection provided by normal ocular flora and predispose to development of opportunistic infections of the cornea.
The use of topical corticosteroids can create a localized immunosuppression and present a major risk factor for bacterial keratitis. Corticosteroids prevent neutrophil migration in response to chemotactic factors released during microbial infection (
77). Impairment in opsonization is a well-known predisposing factor to infection with encapsulated bacteria, including
S. pneumoniae, Streptococcus pyogenes, Haemophilus influenzae, and certain strains of
P. aeruginosa. Premature infants, patients with immunoglobulin deficiencies, asplenic individuals, and patients with sickle cell disease (
78) or systemic lupus erythematosus all may have impaired opsonization.
Passive acquired maternal immunity begins to decline around 3 to 6 months of age before development of actively acquired immunity. This is a relative period of vulnerability for young children to infection with encapsulated bacteria, particularly
H. influenzae (
79). This provides the rationale for immunization with the type B conjugated vaccine against
H. influenzae.Individuals with functionally abnormal leukocytes may be more susceptible to development of marginal keratitis and blepharoconjunctivitis in part because of a loss of control over normal eyelid flora (
80). A variety of systemic conditions can result in compromise, including use of immunosuppressive drugs, extensive body burns, pregnancy, chronic alcoholism, severe malnutrition, immunodeficiency syndromes, including acquired immunodeficiency syndrome (AIDS), drug addiction, malignancy, infancy, and old age (
81). Corneal infection may be facilitated in the systemically compromised host. Patients with AIDS do not appear to have an increased risk for bacterial keratitis, yet may have a more fulminant clinical course (
82,
83). Severe
Pseudomonas keratitis may develop in patients with AIDS, especially in association with contact lens wear (
82).
Because the intact corneal epithelial barrier is such an important first line of defense, development of an epithelial defect represents a major risk factor for infection. Corneal abrasion, foreign body, or erosion may precipitate development of bacterial keratitis. Conditions that result in reduction of the integrity and adherence of the corneal epithelium, including bullous keratopathy, chemical injury, contact lens wear, prior viral infection, especially with herpes simplex virus, and corneal anesthesia (neurotrophic keratopathy), may predispose to development of corneal infection. Toxic epithelial changes from topical medications, including antiviral drugs, antibiotics, or anesthetics, may compromise the host for development of bacterial keratitis. The inhalational use of cocaine may lead to a relative systemic anesthetic state along with hallucinogenic stupor, loss of blink reflex, and development of infectious keratitis (
84). Prescribed topical ocular medications may become contaminated and result in bacterial keratitis, including severe gram-negative infection with
Pseudomonas, Serratia, and
Proteus (
85,
86). Even microtrauma to the corneal epithelium may predispose to subsequent adherence and invasion of microorganisms. Seemingly trivial trauma with contaminated matter, foreign bodies, makeup, or contact lenses, including cosmetic costume plano lenses, can result in organism inoculation (
87).
Contact lens wear of any type has been increasingly recognized as a major principal risk factor for microbial keratitis (
13,
14,
15,
16,
17,
18,
19,
88). Several investigations of contact lens-associated corneal infection have determined a greater risk with extended-wear soft contact lenses and overnight wear compared with daily-wear hard or soft lenses (
89,
90,
91). The risk of ulcerative keratitis was found to be 10 to 15 times greater with extended-wear soft contact lens use than with those lenses worn on a daily basis (
91). Continuous wear may result in cumulative risk because the longer lens wear was extended on a continuous basis, the greater the overall risk. From these early studies, it was estimated that the number of individuals contracting microbial keratitis in the United States because of extended-wear contact lenses is approximately 8000 per year (
90). Smoking may be an additional risk factor for development of ulcerative keratitis among users of soft contact lenses (
91). Extended wear with continuous overnight use promotes tissue hypoxia, as well as decreased tear flow beneath the contact lens, hampering the defensive cleansing function of the preocular tear film and the eyelids. Smoking may also contribute to relative corneal hypoxia, as in other body sites.
The introduction of disposable contact lenses was embraced with considerable enthusiasm in ophthalmology because it was postulated that by eliminating the frequent handling related to daily insertion, removal, and cleaning, these disposable lenses would have a reduced chance of contamination, possibly reducing the frequency of contact lens-associated infection (
92). Use of disposable extended-wear contact lenses, however, does not appear to have significantly decreased the risk (
93,
94,
95,
96). Using contact lenses on a daily-wear basis is relatively safer, but still may result in ulcerative keratitis, with an estimated annual incidence of 3 per 10,000 contact lens wearers (
90). Aphakic contact lens wearers have an estimated six to nine times greater rate of infectious corneal ulceration compared with cosmetic contact lens wearers (
97). Individuals using aphakic contact lenses may be either young or elderly with relative systemic immunosuppression. In addition, the increased risk among aphakic contact lens wearers may be due to prolonged continuous wear with increased corneal hypoxia and epithelial trauma from thick aphakic contact lenses, or to reduced localized defense mechanisms after ocular surgery.
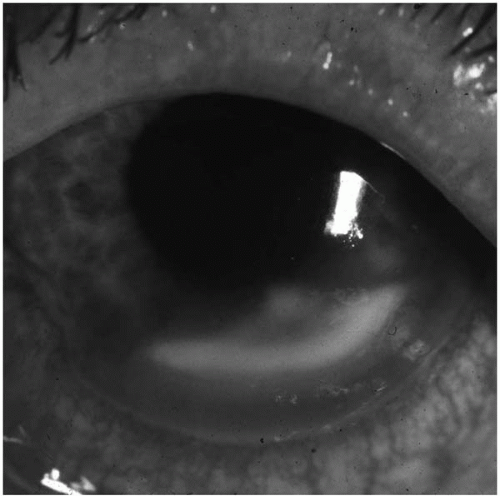
Gram-negative organisms, especially
Pseudomonas, are the most common organisms associated with contact lens-related bacterial keratitis (
19,
88,
98,
99) (
Fig. 13.1-1). With approximately 25 million individuals using contact lenses in the United States, corneal infection is a relatively rare occurrence. Poor contact lens hygiene practices increase the likelihood for development of corneal infection, yet microbial infection may also occur in individuals after proper lens hygiene techniques (
100).
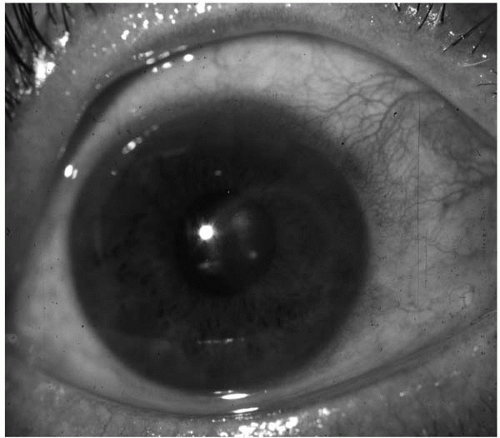
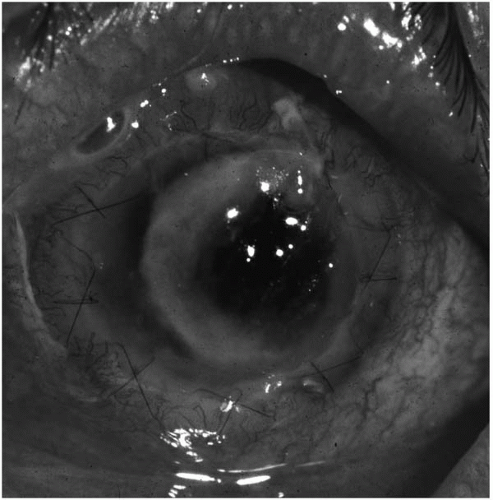
Keratorefractive surgery may introduce potential pathogens deep into the stroma after radial keratotomy (
Fig. 13.1-2) and can result in bilateral, simultaneous keratitis after bilateral surgery (
101,
102,
103). Disruption of the corneal epithelial layer after excimer laser photorefractive keratectomy with or without placement of a therapeutic soft contact lens may be an additional risk factor for the development of bacterial keratitis (
104,
105).
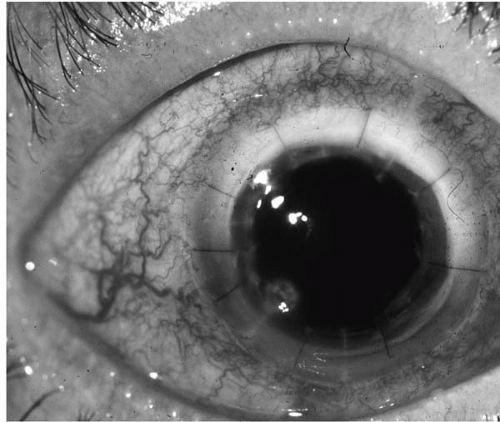
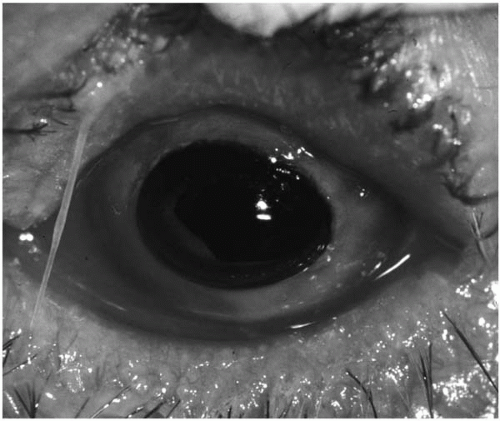
Patients who have undergone penetrating keratoplasty are at increased risk for bacterial keratitis because of adjunctive
chronic topical corticosteroid use, therapeutic soft contact lens placement, and loose sutures (
106,
107,
108) (
Fig. 13.1-3).
Traumatic and systemic disease may be frequently associated and increase the risk for development of bacterial keratitis in children (
109,
110,
111). Before 3 years of age,
Pseudomonas was the most common organism recovered, whereas in older children,
S. aureus, S. pneumoniae, and
P. aeruginosa were the most common causative organisms in a series of children with bacterial keratitis (
110). Because of potential delay in diagnosis and difficulty in the delivery of therapy, surgical intervention may be required (
110,
111).
A retrospective analysis of hospital records of patients with bacterial keratitis in Paris, France was conducted to identify predisposing factors and to define clinical and microbiologic characteristics of bacterial keratitis in current practice (
112). The study included 300 cases (291 patients) of presumed bacterial keratitis. Potential predisposing factors, usually multiple, were identified in 90.6% of cases. Contact lens wear was the main risk factor (50.3%), whereas trauma or a history of keratopathy was found in 15% and 21% of cases, respectively. An organism was identified in 201 eyes (68%). Eighty-three percent of the infections involved gram-positive bacteria, 17% involved gram-negative bacteria, and 2% were polymicrobial. Gram-negative bacteria were associated with severe anterior chamber inflammation, as well as greater surface of infiltrates. Ninety-nine percent of ulcers resolved with treatment. However, only 60% of patients had visual acuity better than the level at admission, and 5% had a very poor visual outcome (
112).
Orthokeratology is a process by which the corneal curvature is flattened by sequentially fitting rigid, gas-permeable contact lenses of decreasing central curvature. There has been a resurgence of interest in the procedure with the recent introduction of reverse-geometry lenses. Although promising results have been described in reducing the myopic refractive error, the use of these lenses can be associated with corneal problems, as reported in an observational case series that identified six children with orthokeratology-related corneal ulcers (
113).
Consecutive cases of orthokeratology lens (OKL)-related corneal ulcers in children presenting to a tertiary
referral center (March 1999 to June 2001) were reviewed for preinfection and postinfection visual acuity, refraction, and any organisms identified. Six children between the ages of 9 and 14 years (mean, 12.1 years) were treated. The male-female ratio was 1:5. All cases were unilateral, with equal numbers of left and right eyes. All children wore the OKL at night for 8 to 12 hours, with the onset of infection between 3 and 36 months (mean, 16.6 months) of OKL wear. All of the patients suffered a resultant BCVA loss. Five of the six cases were culture positive for
P. aeruginosa (
113). In view of the temporary benefits of orthokeratology, together with a known increased risk of infection associated with overnight lens wear, parents of children considering orthokeratology must be informed and warned of the potential for permanent loss of vision. The ophthalmic community should have a heightened awareness of the associated complications.
PATHOGENESIS
The pathogenesis of ocular infectious disease is determined by the intrinsic virulence of the microorganism, the nature of the host response, and the anatomic features of the site of the infection (
114). The avascular, clear anatomic structure of the cornea with its specialized microenvironment predisposes to potential alteration and destruction by invading microorganisms, virulence factors, and host response factors. The intrinsic virulence of an organism relates to its ability to invade tissue, resist host defense mechanisms, and produce tissue damage (
115). Penetration of exogenous bacteria into the corneal epithelium typically requires a defect in the surface of the squamous epithelial layer. By virtue of specialized enzymes and virulence factors, a few bacteria, such as
N. gonorrhoeae, N. meningitidis, C. diphtheriae, Shigella, and
Listeria, may directly penetrate corneal epithelium to initiate stromal suppuration.
Bacteria colonize host cells by engaging adhesins at their surface with receptors on the host cell surface. Specific receptors are often required by many adhesins to achieve binding. Besides adherence, microbial adhesins also contribute to subsequent interactions. Virulence factors may initiate microbial invasion or secondary effector molecules may assist the infective process. Upregulation or downregulation of host defense mechanisms may be involved. Adhesins may also be toxins (
116). Therefore, receptor recognition is only the first step in the pathogenesis of infection directed by microbial adhesin molecules (
117).
Many bacteria display several adhesins on fimbriae (pili) and nonfimbriae structures. Such adhesive proteins may recognize carbohydrates on host cells; alternatively, protein-protein interactions can also occur.
Certain bacteria exhibit differential adherence to corneal epithelium. The adherence of
S. aureus, S. pneumoniae, and
P. aeruginosa to ulcerated corneal epithelium is significantly higher than that of other bacteria and may account in part for their frequent isolation (
118).
P. aeruginosa has many virulence factors that contribute to pathogenesis. Cell-associated structures such as pili (
119) and flagella (
120), and extracellular products, such as alkaline protease (
121), elastase (
121), exoenzyme S (
116), exotoxin A (
122), endotoxin (
123), slime polysaccharide (
124), phospholipase C (
121), and leukocidin (
121), are associated with virulence, invasiveness, and colonization. Whereas gram-positive bacteria, including
S. aureus, adhere to host tissues through fibronectin and collagen (
125),
P. aeruginosa attaches to cell surfaces that lack fibronectin (
126). Bacteria adhere to injured cornea (
127), to exposed corneal stroma (
128), or to immature nonwounded cornea (
129). The corneal epithelial receptors for
Pseudomonas species are glycoproteins (
130,
131).
Pili (fimbriae) are thin (4 to 10 nm in diameter) protein filaments that are located on the surfaces of many bacteria.
In vitro studies indicate purified pili successfully compete for binding with cold bacteria by saturating available binding sites on the ocular surface (
132). Monoclonal antibodies specific for
Pseudomonas pili and a peptide-conjugated alkaline phosphatase allowed identification of host corneal receptor molecules (
133). Characterization of these receptor proteins indicates that carbohydrates are necessary for receptor activity (
131). In competitive inhibition experiments, sialic acid was the only aminosugar able completely to inhibit pilus binding to mouse corneal epithelial proteins. Pili have been used to protect against
P. aeruginosa keratitis (
132). Pili, however, have considerable antigenic variation between strains (
134). Bacteria also possess an array of other virulence factors, including non-pilus adhesins (
135). Some clinical isolates of
P. aeruginosa from keratitis are reportedly nonpiliated (
136).
Flagella are subcellular filamentous organelles (16 to 18 nm in diameter) originating in the cell membrane and extending to 15 to 20
μm from its surface. These organelles are responsible for bacterial motility, which is important for dissemination of infection (
137). The flagellum has a helicofilament composed of cell-assembling subunits of the protein flagellin. More than 95% of
P. aeruginosa clinical isolates are flagellated (
138). Because flagella contain aminospecific proteins and have a large surface exposure available for antibody binding, they have been considered as the basis for vaccine development. Much research examining the flagellum as a virulence factor in
P. aeruginosa infection or as a vaccine candidate has focused on the burned-mouse model (
139). A loss of virulence was observed in flagella-deficient mutants active or passive systemic, as well as topical immunization with flagella or topical antiflagellar antibody homologous to the infecting strain of bacteria, protects mice against
P. aeruginosa corneal infection (
140).
In addition to adhesins, the adherence of
P. aeruginosa and
N. gonorrhoeae is guided by glycocalyx, a biologic slime that enables them to adhere to susceptible cells, producing slime aggregates that are resistant to phagocytosis (
141). Similar coatings may form on contact lenses to facilitate the adherence of bacteria to the lens material (
142).
After adherence of microorganism to the cornea and initiation of infection, the complex tissue reactions of the host response occur, including inflammation, neovascularization, cellular and humoral immune responses, and stromal degradative processes.
A variety of cytokines may be released in response to corneal infection. During inflammation, leukocyte adhesion to the vascular endothelium is enhanced by interleukin-1 (IL-1) and tumor necrosis factor (TNF), products of macrophages and T lymphocytes (
143). IL-1 is a known potent intracellular mediator of inflammation and chemotaxis for neutrophils. TNF stimulates immunocompetent cells and induces release of IL-1 and IL-6 from macrophages and other cells (
144). Bacterial exotoxin-A downregulates TNF, IL-1, and lymphotoxin by inhibiting the cells’ ability to produce these cytokines (
145).
Neutrophil infiltration after corneal infection is a principal host defense mechanism. The host inflammatory response to
Pseudomonas species has been studied in mice designated as susceptible or resistant. Susceptible mice are unable to clear the cornea, whereas resistant mice may restore corneal clarity within 1 month after ocular challenge (
146). Resistant mice appear to have a larger number of corneal leukocytes present initially. The resistant mice also have a shorter duration of inflammatory cell and bacterial residence in the cornea compared with susceptible mice.
If neutrophils are experimentally depleted, many resistant animals succumb to lethal
Pseudomonas sepsis within 48 hours (
147). In aged mice, deficiencies of neutrophilic phagocytosis have been observed. These observations may partially explain the clinically apparent age-related susceptibility of individuals to corneal infection.
Preimmunization of rats with phenol-killed
Pseudomonas organisms still results in massive corneal stromal degradation caused by neutrophil migration in the absence of viable bacteria. Naive, unimmunized mice show little stromal destruction during early infection, despite the presence of numerous bacteria in the cornea. Thus, immune recognition is involved in the host response to
Pseudomonas corneal infection and is apparently required for phagocytosis, but not for neutrophil recruitment (
148).
Models of experimental
P. aeruginosa ocular disease also indicate the importance of the third component of complement (C3) in host resistance to corneal infection. Resistant mice experimentally depleted of C3 by cobra venom and then inoculated with
Pseudomonas organisms to the cornea respond with delayed leukocyte mobility, bacterial persistence in the cornea, and subsequent scarring and opacification (
149).
In addition to organism factors, host lysosomal enzymes and oxidative substances produced by neutrophils, keratocytes, and epithelial cells may significantly contribute to the destruction caused by
Pseudomonas keratitis (
150). Two eukaryotic gelatinase species have been characterized, including a type IV collagenase (72 kD) and a type V collagenase (90 to 100 kD) (
151). Corneal epithelial cells produce predominantly the 92-kD form of progelatinase, whereas stromal fibroblasts synthesize predominantly the 72-kD progelatinase (
152). The progelatinase is cleaved by
Pseudomonas elastase to produce the active form (
153).
A variety of bacterial toxins and enzymes may be produced during corneal infection to contribute to destruction
of the corneal substance.
P. aeruginosa produce many toxic substances capable of causing necrotic central corneal ulceration. Toxin-A inhibits protein synthesis much as diphtheria toxin by catalyzing the transfer of the adenosine 5′-diphosphate ribose (ADPR) portion of nicotinamide adenine dinucleotide to mammalian elongation factor II. With staphylococcal keratitis, alpha-toxin, but not protein A, is a major virulence factor mediating corneal destruction (
154). Advances in our understanding of host innate and adaptive immune responses to experimental infection with
P. aeruginosa have been made using a variety of animal models, including inbred murine models that are classed as resistant (cornea heals) versus susceptible (cornea perforates) (
155). Evidence has been provided that sustained IL-12-driven interferon (IFN)-
γ production in dominant T-helper-1 responder strains such as C57BL/6 (B6) contributes to corneal destruction and perforation, whereas IL-18-driven production of IFN-
γ in the absence of IL-12 is associated with bacterial killing and less corneal destruction in dominant T-helper-2 responder strains such as BALB/c. The critical role of IL-1 and chemotactic cytokines such as macrophage inflammatory protein-2 in PMN recruitment and the critical role of this cell in the innate immune response to bacterial infection is reviewed (
155). Regulation of PMN persistence is also discussed, and evidence provided that persistence of PMN in B6 cornea is regulated by CD4
+ T cells, whereas macrophages regulate PMN number in the cornea of BALB/c mice (
155). The studies provide a better understanding of the inflammatory mechanisms that are operative in the cornea after
P. aeruginosa challenge and are consistent with long-term goals of providing targets for alternative or adjunctive treatment for this disease. Future studies will be aimed at better defining the role of Toll receptors, neuropeptides (as unconventional modulators of the immune response), and exploitation of disease control by new techniques, such as RNA silencing (
155).
In summary, the pathogenesis of bacterial keratitis initially requires the adherence of bacteria to disrupted or normal corneal epithelium. Adhesins are microbial proteins that direct the high-affinity binding to specific cell surface components. These adhesins are able to promote bacterial entry into the host cell, derange leukocyte migration, activate plasmin, and induce cytokine production. In addition, they may act as toxins directly. Adhesins recognize carbohydrate and protein moieties on the host cell surface. Most bacteria can display a number of adhesins. Although the cognate oligosaccharides for bacterial adhesins are known, the molecules bearing these determinants are not well characterized. Integrins are a family of glycoproteins mediating cell-cell and cell-extracellular matrix recognition. Many bacterial pathogens have coopted the existing integrin-based system, masking ancillary ligand recognition in a form of mimicry. Once the bacterial pathogen has adhered to the corneal epithelial surface, the next step in establishing infection is invasion into the corneal stroma. Bacterial invasion is facilitated by proteinases that degrade basement membrane and extracellular matrix and cause cell lysis. Proteinases may be derived from bacteria, corneal cells, and migrating leukocytes. Corneal matrix metalloproteinases (MMPs) are excreted in an inactive form, but are activated during infection by bacterial proteinase. Corneal proteinase production may also be induced during the course of infection. The invasion of bacteria into the cornea is facilitated by a number of exotoxins, including P. aeruginosa phospholipase, heat-stable hemolysin, and exotoxin-A, which leads to stromal necrosis. Once bacterial invasion into the cornea has ensued, infection is further facilitated by a complex sequence leading to interruption of the host immune response. Exopolysaccharide formation by both gram-positive and gram-negative bacteria results in local immunosuppressive effects. Certain bacteria with capsular polysaccharide also have immunosuppressive properties, including interference with phagocytosis. Proteases degrade complement components, immunoglobulins, and cytokines and may inhibit leukocyte chemotaxis and lymphocyte function. Toxin-A inhibits protein synthesis much as diphtheria toxin by catalyzing the transfer of the ADPR portion of nicotinamide adenine dinucleotide to mammalian elongation factor-II. Exoenzyme-S is another ADP-ribosyl transferase that may act as an adhesin and also contribute to dissemination of the organism.
Two specific bacterial proteases, elastase and alkaline protease, cause marked destruction of the cornea when injected intrastromally (
156,
157). Intrastromal injection of purified elastase alone also results in severe corneal damage. Inhibition of elastase activity with 2-mercaptoacetyl-L-phenylalanine-L-leucine (
158) prevents keratolysis. The proteases contribute to the pathogenesis of keratitis by degrading basement membrane (
159), laminin, proteoglycans, extracellular matrix (
160), and collagen (
161). In addition, the bacterial proteases inhibit host defense systems by degrading immunoglobulins, IFN, complement, IL-1, IL-2, and TNF (
156). Such interference results in decreased neutrophil chemotaxis, T-lymphocyte function, and natural killer cell function. Mutants deficient for alkaline protease do not establish corneal infection, suggesting that this protease is an important initiating factor (
162). A bacterial heat-labile phospholipase C has been shown in antibody/substrate specificity studies to be produced in mouse ocular infections, suggesting its role as a potential virulence factor (
163). Bacterial lipopolysaccharide stimulates neutrophil migration and infiltration into the cornea with subsequent corneal scarring and opacification (
164).
Bacterial exotoxins are released by actively replicating organisms, and some endotoxins are released only after the death of the organism. These enzymes and toxins have been shown to persist in the cornea for a protracted period and continue to cause stromal destruction after the death of the
pathogen. Most of the bacterial exotoxins are thermal labile and have antigenic properties. Gram-positive bacteria elaborate a variety of biologically active and immunologically distinct toxins. Coagulase-positive strains of staphylococci are the most pathogenic, and elaborate other extracellular enzymes, such as staphylokinase, lipase, hyaluronidase, DNase, coagulase, and lysozyme. Coagulase-negative staphylococci, including
S. epidermidis, also produce potentially destructive toxin (
165). Streptococcal toxins include streptolysin O and S, erythrogenic toxin, and the enzymes hyaluronidase, streptodornase, and streptokinase. The invasiveness of
S. pneumoniae is aided by collagenase activity (
166), although the organism may be inherently invasive without toxin production.
The lipopolysaccharides composing the endotoxins in the cell wall of gram-negative bacteria may be released on the death of the organism. These lipopolysaccharides may result in the production of stromal rings (
Fig. 13.1-4). These rings have been shown to consist of polymorphonuclear leukocytes in the corneal stroma, which have been chemoattracted by the alternative complement pathway (
167,
168). In addition to gram-negative bacterial keratitis, such ring infiltrates have been described in fungal, viral, and
Acanthamoeba keratitis. In nonbacterial corneal infection, these stromal rings are thought to result from antigen-antibody precipitants (immune rings).
CLINICAL FEATURES
Once corneal infection is established, there are no absolutely specific clinical symptoms to confirm infection or exclusively distinctive biomicroscopic signs to distinguish the responsible organism(s). The determinants of clinical presentation include the strain (virulence) of the responsible organism(s), the method of inoculation or introduction, the time interval from inoculation, the antecedent status of the cornea, prior or concomitant antimicrobial corticosteroid therapy, and other host factors (
115).
Because of the rich innervation of the cornea, the most common symptom of inflammatory lesions of the cornea is pain. Movement of the eyelids over ulcerated corneal epithelium intensifies the pain. Therefore, examination of patients with suspected microbial keratitis is greatly facilitated by instillation of topical anesthetic. Keratitis is usually accompanied by a variable decrease in vision. Reflex tearing, photophobia, and blepharospasm are common. A discharge, which is a distinctive feature of conjunctivitis, is usually absent in patients with keratitis, unless an associated purulent conjunctivitis is present, such as with gonococcal, pneumococcal, and Haemophilus infections. The conjunctiva may be variably hyperemic and a nonspecific papillary reaction may vary in intensity, depending on the severity of the keratitis. The preocular tear film in bacterial keratitis can be observed by slit-lamp microscopy to contain inflammatory cells and debris. Ipsilateral lid edema may be variably observed with bacterial keratitis. In addition to conjunctival hyperemia, chemosis may develop, especially with highly virulent causative organisms.
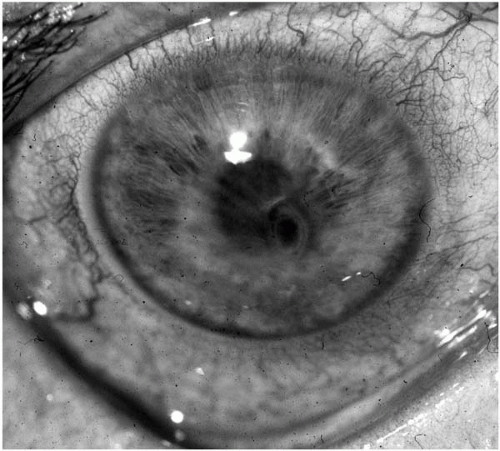
The hallmark clinical signs that are distinctive for suspected infectious keratitis include an ulceration of the epithelium with suppurative stromal inflammation that is either focal or diffuse (
Fig. 13.1-5). Multifocal suppurative inflammation in the cornea is suggestive of mixed infection (polymicrobial keratitis) (
169). Polymicrobial keratitis has
been observed in from 6% to 56% of overall cases (
170,
171). Microbial keratitis may occasionally present with an intact epithelium and nonsuppurative multifocal stromal inflammation. The presence of diffuse cellular infiltration in the adjacent stroma and an anterior chamber cellular reaction is highly suggestive for infectious keratitis. The anterior chamber reaction may range from mild flare and cells to severe layered hypopyon formation.
The hypopyon in bacterial keratitis is usually sterile when Descemet’s membrane is intact. Hypopyon may also be present with mycotic keratitis and severe necrotizing viral stromal keratitis. Certain noninfectious conditions may be accompanied by hypopyon formation, including Behçet’s syndrome, severe alkali injury, therapeutic soft contact lens use, and abuse of topical anesthetic agents.
If the antecedent status of the cornea is abnormal, the clinical signs and symptoms may be nondistinctive. Interpretation of the clinical signs may be difficult if there has been prior corneal inflammation, uveitis, and structural abnormalities. Careful monitoring of symptoms of pain and redness, as well as increasing epithelial or stromal ulceration, increasing anterior chamber inflammatory reaction, or an overall acute deterioration in the clinical status, should elevate the index of suspicion for infection (
172). Prior treatment with antibiotic therapy can blunt the clinical symptoms and signs of corneal infection. Prior treatment with corticosteroids may enhance the likelihood of invasion by opportunistic organisms and dampen the clinical signs of inflammation associated with bacterial keratitis. Corticosteroids and other immunosuppressive agents may retard host defense mechanisms and inhibit chemotaxis, neutrophil migration, phagocytosis, degranulation, and lysosomal enzyme release. The use of corticosteroids before infection may also result in severe rebound inflammation in the stroma on abrupt cessation of corticosteroid therapy during the course of infection. Such a rebound inflammation may confound the clinician’s ability to monitor the initial response to therapy. Surreptitious use of over-the-counter vasoconstrictor medicines may mask conjunctival hyperemia. The marked corneal toxicity of particular agents, including aminoglycosides, topical anesthetics, topical antivirals, and topical amphotericin B, can mimic bacterial keratitis by causing epithelial and stromal ulceration, stromal infiltration, and anterior chamber inflammatory reaction. Prior contact lens wear can also alter the clinical presentation of bacterial keratitis. Infection associated with contact lenses may be multifocal with diffuse epithelial and stromal infiltration. Given the ability of bacteria to bind to contact lenses, individuals presenting with corneal abrasions in association with contact lens wear should be suspected of having possible early bacterial keratitis. Placing a patch over a corneal epithelial abrasion associated with contact lens wear may create conditions favorable for the development of infectious keratitis (
173).
Prompt clinical recognition of bacterial corneal infection is essential to successful management. The patient’s symptoms and clinical biomicroscopic signs alone may be insufficient reliably to diagnose bacterial keratitis. In a study of patients hospitalized for ulcerative keratitis, diagnostic evaluations disclosed that 65% of the patients had nonbacterial keratitis (
14). The differential diagnosis of bacterial keratitis may be particularly difficult in cases of herpes simplex viral keratitis, neurotrophic ulceration, marginal ulcerative and infiltrative keratitis from multiple causes, toxic keratopathy, and those diseases characterized by persistent epithelial or anterior stromal ulceration. With herpes, a prior history of herpetic infection and a relative asymmetrically diminished corneal sensation may be helpful to distinguish herpes simplex viral from bacterial keratitis. Patients with keratoconjunctivitis sicca may be especially sensitive to the toxic effects of topical medication, which can result in keratolysis and perforation in the absence of stromal cellular infiltration.
A clinical assessment of the severity of the bacterial keratitis should be made at the initial presentation. Careful measurement and documentation of objective parameters for comparative analysis with subsequent remeasurements are important to monitor the clinical course. Using the adjusting slit beam on the biomicroscope, the overall size of the epithelial involvement can be measured by recording the diameter in two dimensions. Similarly, the area of stromal ulceration can be measured in two meridians. An estimate of the depth of stromal ulceration should be determined by comparing adjacent uninvolved corneal thickness. Slit-lamp photographs are helpful for documentation and monitoring of the clinical course. Initial corneal topographic analysis may be helpful in select cases. Detailed clinical drawings with measurements of the size and depth of infiltration should be recorded at each visit. Additional features to assess include the intensity of suppuration and edema, thickness of the stroma, accompanying scleral suppuration, the degree of anterior chamber and iris inflammation, secondary glaucoma, and the rate of progression or pace of inflammation. A grading system based on the characteristics, including the size of the ulceration in millimeters, percentage depth of ulceration, intensity or density of infiltration, and scleral involvement may provide a guide to the aggressiveness of therapy. More detailed grading systems have been described (
174).
Certain characteristic clinical features may be suggestive of specific corneal pathogens, although clinical observation alone should not replace laboratory investigation with corneal scrapings for smears and culture (
172,
175,
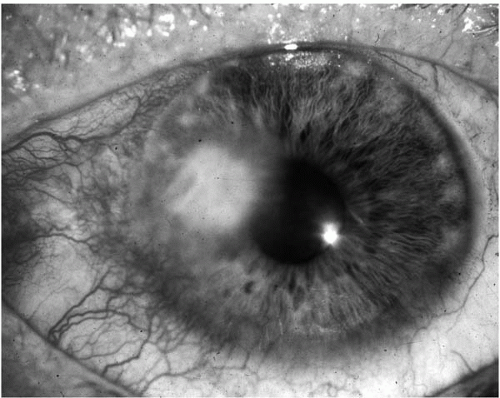
176). Gram-positive cocci typically cause localized, round or oval ulcerations with grayish-white stromal infiltrates having distinct borders and minimal surrounding epithelial edema (
Fig. 13.1-6). Staphylococcal keratitis is more frequently encountered in compromised corneas, such as with bullous keratopathy, chronic herpetic keratitis, keratoconjunctivitis sicca, ocular rosacea, or atopic keratoconjunctivitis. With
delay in presentation and long-standing infection, both coagulase-positive and coagulase-negative staphylococcal keratitis may cause severe intrastromal abscess and corneal perforation.
Staphylococcus is a common inhabitant of the normal ocular flora. Coagulase-negative staphylococci may be recovered from eyelid cultures in over 85% of healthy subjects, whereas
S. aureus has been documented in 15% of eyelid cultures (
177). Excessive colonization of the anterior lid margin with staphylococci may be associated with hypersensitivity (type III), marginal infiltrative keratitis, or peripheral ulceration. In compromised corneas, superinfections with
S. aureus may present with an atypical appearance. In these abnormal corneas,
S. aureus infection may result in marked suppuration with a deep stromal abscess. An accompanying large hypopyon or endothelial fibrin plaque is usually present and larger than anticipated, based on the area of ulceration.
S. epidermidis typically has a more indolent clinical course, yet may also lead to deep stromal abscess and the potential for corneal perforation.
After trauma,
S. pneumoniae keratitis may present with a deep, oval, central stromal ulceration having serpiginous edges (
Fig. 13.1-7). There is typically dense stromal abscess formation with radiating folds in Descemet’s membrane and moderate accompanying stromal edema. Hypopyon with retrocorneal fibrin deposition is a common clinical feature. Progression to corneal perforation is possible. Some strains of
S. pneumoniae (designated S strains) are encapsulated, whereas others (the R strains) are not. Virulence for corneal infection is not totally dependent on encapsulation because R strains have been recovered from clinical keratitis (
178). An abnormal antecedent cornea may modify the classic serpiginous, hypopyon ulcer described after trauma with
S. pneumoniae infection. Beta-hemolytic streptococci may cause severe corneal infection with dense suppuration, which may progress to perforation. A distinct, indolent, pauciinflammatory-appearing crystalline keratopathy has been observed in association with streptococcal corneal infection (
179,
180,
181,
182,
183).
Gram-negative corneal infection typically follows a rapid-paced inflammatory destructive course or, alternatively, a less commonly encountered, slowly progressive
indolent ulceration.
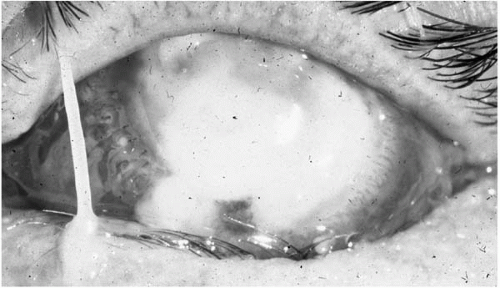
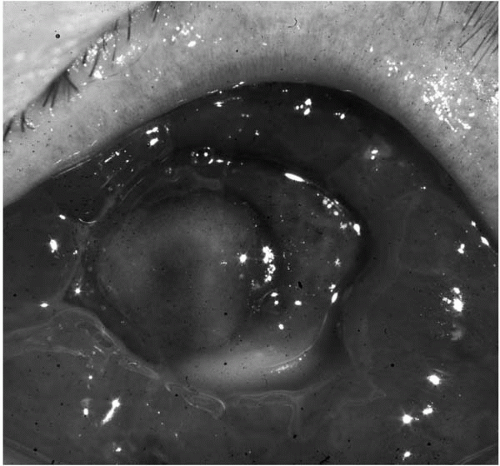
P. aeruginosa has the most distinctive clinical course after corneal infection. There is a loss of corneal transparency with adjacent peripheral inflammatory epithelial edema and a “ground-glass” stromal appearance. The keratitis may progress rapidly, even with appropriate treatment, into a deep stromal abscess (
Fig. 13.1-8) spreading concentrically to form a ring ulcer with large hypopyon (
Fig. 13.1-9). Complete stromal keratolysis with perforation may occur, even after apparent clinical improvement.
A less common clinical presentation of
Pseudomonas keratitis behaves in a more indolent fashion and the organism may not possess similar virulence factors, including proteoglycanolytic enzymes, as discussed in the pathogenesis section. Other gram-negative rods may cause slowerpaced ulcerative keratitis in previously compromised corneas with less intense anterior chamber inflammatory reaction and less specific distinguishing features.
Proteus may have a fulminant course similar to
Pseudomonas without trauma.
Klebsiella is more often associated with infection as a consequence of chronic epithelial disease.
S. marcescens may be an opportunistic pathogen causing contact lens-associated keratitis (
36).
Moraxella was originally discovered as an ocular pathogen.
Moraxella may colonize the nasopharynx and produce keratitis after trauma in debilitated, alcoholic, malnourished, or diabetic patients. The large, boxlike diplobacilli and pleomorphic gram-negative rods of non-
liquefaciens Moraxella may be observed in corneal scrapings from nonalcoholic patients in the presence of a preexisting epithelial defect (
23). The typical clinical features of
Moraxella keratitis include an indolent corneal ulceration with mild to moderate anterior chamber reaction. The ulceration is usually oval with a predilection for the inferior portion of the cornea (
Fig. 13.1-10). In some cases of
Moraxella keratitis, moderately severe stromal and anterior chamber reactions may develop, with endothelial decompensation and possible perforation (
184).
A rapidly paced, hyperpurulent conjunctivitis with marked hyperemia, chemosis, and corneal epithelial ulceration with
stromal infiltration should suggest infection with
N. gonorrhoeae, or
N. meningitidis (
Fig. 13.1-11). A rapid and devastating keratitis may also follow trauma and contamination with
B. cereus. B. cereus keratitis corneal infection is characterized by a distinctive stromal ring infiltrate remote from the site of injury with rapid progression to stromal abscess, corneal perforation, and intraocular extension with destruction mediated by specific exotoxins (
32). The presence of a distinctive air bubble in the anterior chamber or in the corneal stromal beneath the epithelium, especially after trauma with contaminated soil, should suggest possible infection with spore-forming
Clostridium species (
42).
Corneal infections with
Nocardia, Actinomyces, and
Streptomyces typically follow an indolent clinical course, which may simulate mycotic keratitis with hyphal edges, satellite lesions, and elevated epithelial lesions. A chronic epithelial defect with “calcareous” bodies at the edges of epithelial ulceration was caused by
Nocardia asteroides (
185).
The vast majority of ocular infections by nontuberculous mycobacterial species involve the cornea and are principally due to the rapid growers,
M. fortuitum and
M. chelonae (
186).
M. fortuitum has been recognized as an opportunistic pathogen capable of causing intractable keratitis frequently resistant to multiple antibiotics (
49). Between 1965 and 1974, virtually all of the reported cases of keratitis secondary to nontuberculous mycobacteria were attributed to
M. fortuitum. In more recent years (1978-1995), there has been a preponderance of
M. chelonae isolates accounting for the majority of reported corneal infections (
187,
188,
189,
190,
191). Clinically, the nontuberculous mycobacteria produce relatively slowly progressive keratitis, which may mimic the course of disease due to other indolent organisms, such as fungi or anaerobic bacteria. Environmental exposure is the most common route of inoculation and many patients have a history of antecedent trauma or prior surgery, such as penetrating keratoplasty or radial keratotomy. Clinical infection usually develops 2 to 8 weeks after the corneal trauma; however, delayed-onset keratitis occurring 2 years after radial keratotomy has been reported (
191). Treatment with topical corticosteroids before the identification of the infection frequently occurs. Experimental studies have suggested that
M. fortuitum keratitis in rabbits is made worse by corticosteroid use (
192). The typical clinical features of nontuberculous mycobacterial keratitis include a relative paucity of suppuration, although dense stromal abscesses may form and rarely corneal perforation may occur. Multifocal lesions may be observed at the time of presentation and satellite lesions may also develop as the infection worsens. The presenting corneal lesion has occasionally been linear and pseudodendritiform, with accompanying epithelial ulceration. In partially treated cases, the lack of response to conventional antibiotic therapy is frequently a clue to the diagnosis. In addition to the more typical clinical features, a broad spectrum of unusual presentations has been reported, including ring stromal infiltrate (
193) and crystalline keratopathy (
194).