More than 35 million people in the United States are aged 65 years or older. Of people aged more than 80 years, 62% to 80% have a significant reduction in their sense of smell, which can adversely affect their safety and nutritional status. This article reviews the anatomy of smell and discusses the known and theorized etiologies of smell loss in the elderly population.
At least 35 million people aged 65 years or older are living in the United States, accounting for 13% of the US population. This number is estimated to increase to 75 million by 2030. Although not a life-threatening problem, the loss of taste and smell can result in significant changes in appetite and food preferences and, in turn, affect the quality of life and nutritional status in the elderly population. Intact smell is an important warning system that alerts us to the presence of leaking gas, smoke, food spoilage, and pollution. The absolute sensitivity to odor and the appreciation of suprathreshold odors decline with increasing age. In many elderly patients there is unawareness of actual smell loss. Nardin and colleagues showed that 77% of older patients with smell loss reported normal smell sensitivity. Smell sensation deficits occur in many neurodegenerative diseases such as Alzheimer’s and Parkinson’s disease which typically affect the elderly. The etiology of smell loss in older patients has yet to be completely defined. The loss of smell sensitivity can be due to conditions that could affect any step in the olfactory process from the transport of odors up to the olfactory cleft to the central processing of olfactory information.
Anatomy and physiology of the sense of smell
Odors reach the olfactory epithelium orthonasally via the nose or retronasally via the oropharnx. Retronasal function is essential for the proper appreciation of flavor of food. Odorants are detected when they bind to receptor proteins present in the olfactory receptor neurons which are located in the olfactory epithelium in the dorsal aspect of the nasal cavity ( Fig. 1 ). These receptor cells have nonmotile ciliary projections that express one of several hundred different olfactory receptor proteins. The olfactory receptor function is dependent on the composition of the mucous layer that is produced by the Bowman’s glands and sustentacular cells. This mucous layer contains mucopolysaccharides, enzymes, and immune factors which can help modify olfactory stimulants but also have a role in the detoxification of inhaled substances, the protection of other organs, and the activation of toxicants. The composition of nasal secretions can change dramatically with inflammation, toxic exposure, or disease. Because the ciliary projections of the olfactory receptor neurons are nonmotile, mucociliary clearance of the olfactory cleft is solely dependent on the ciliary function of the respiratory epithelium. Normal respiratory ciliary function can be reduced by toxic exposures such as cigarette smoke.
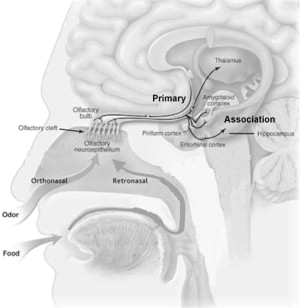
Activation of an olfactory receptor neuron results in G-protein CAMP cascade, which leads to cell depolarization. Olfactory receptor neurons expressing the same receptor proteins converge onto the same loci in the olfactory bulb. This process ensures transfer of a given odorant signal to a specific set of mitral cells in the olfactory bulb. The signal can then propagate centrally via the olfactory tracts and the lateral olfactory striae and terminate in the amygdala and the primary olfactory cortex of the uncus. Projections can then connect to the association olfactory cortex of the parahippocampal gyrus and the entorhinal. The parahippocampal gyrus receives fibers from the cingulate gyrus via the cingulum and projects into the hippocampus. These primary and associative areas are referred to as the pyriform cortex (see Fig. 1 ). The amygdala receives connections from the middle and inferior temporal gyri important for semantic processing. Damage to the pyriform cortex could impair smell at the perceptual or sensory level. Damage to the temporal cortical-amygdala connections could impair associative capabilities.
Mammalian olfactory epithelium maintains the ability to replace olfactory neurons lost via injury. Olfactory neurogenesis, stimulated by the death of olfactory receptor neurons, begins with the maturation of progenitor cells. These precursor cells mature and extend axons through the cribriform plate which synapse within the olfactory bulb where they derive trophic factors needed for long-term survival. Olfactory receptor cells are bipolar neurons directly exposed to the external environment and the dangers of that environment, including injury from infection, inflammatory, and noxious chemical agents. This environmental exposure results in a regular turnover of olfactory receptor cells. Cell death frequently occurs via the process of apoptosis. Apoptosis is a programmed active process that usually begins with the receipt of a death signal that then activates intrinsic enzymes which ultimately lead to cell death. This mechanism is used for the removal of aged or damaged cells, immune regulation, and cancer surveillance and in response to injury. Recent studies by Kern and colleagues have demonstrated increases in levels of capsase 3, the dominant executioner enzyme in the apoptotic pathway, in the olfactory epithelial cells of patients with nasal sinus disease as well as in mice models of traumatic smell loss. This finding suggests an increase in programmed cell death in these conditions.
Smell loss with aging
Many studies have shown a decrease in olfactory ability in humans with advancing age. Researchers have documented this loss using several different psychophysical measures, including odor detection (threshold), quality discrimination, and odor identification. The detection of suprathreshold odors also appears to decrease with age. These decreases in olfactory ability are noted in both sexes after the age of 69 years; however, women are better at identifying odors than men at all ages. After the age of 80 years, 62% to 80% of subjects tested showed major olfactory impairment, with 50% of subjects completely anosmic.
Mucous Layer
The ability to perceive odors may be modified by changes in the mucous layer in the olfactory cleft that occurs with increasing age. The mucous layer influences odor deposition and clearance rates. Studies have shown a decline in hydration and secretion of mucus in the elderly population.
Modification of the olfactory xenobiotic metabolism may have a role in the decline of smell sensitivity over the years that the mucosa is exposed to the environment. Many common environmental exposures may alter nasal enzymatic activity, which could enhance individual variations in responses to toxic exposures. These alterations can be due to direct exposure to chemicals that induce or inhibit nasal enzymes or from toxic insults that may alter the olfactory mucosa. Exposure to heavy metals such as manganese can lead to smell loss that can occur at a subtle degree, often leaving the patient unaware of any impairment, similar to the symptoms seen in patients with Alzheimer’s and Parkinson’s disease. Toxic exposures over a lifetime could have a role in age-related smell loss. Cigarette smoke is another pollutant that can alter olfaction. Rhondase, an enzyme responsible for cyanide metabolism, shows a 50% reduction in activity in smokers when compared with nonsmokers. Some odorous stimuli are potentially harmful to the olfactory mucosa if they reach high enough concentrations and remain in the olfactory cleft for prolonged periods of time. Volatile substances with potentially harmful effects usually are presented to the olfactory epithelium for short periods and in low enough concentrations to avoid causing harm; however, the reduced mucociliary movement, reduced hydration, thinner epithelium, and decreased enzymatic activity seen in older olfactory epithelium may allow these substances to contribute to epithelial damage.
Changes in Olfactory Epithelium
Histologic studies show a replacement of olfactory epithelium with respiratory epithelium during aging ( Fig. 2 ). Animal studies have shown a thinning of the olfactory mucosa with age, a decrease in the number of olfactory neurons present, and an increase in the irregularity of the respiratory/olfactory mucosal border. It appears there may be an increase in natural cell turnover via the apoptotic pathway in older mice. Robinson and colleagues reported increased expression of the procapsase 3 gene in older rats. This gene leads to production of capsase 3, the major executioner enzyme in the apoptotic pathway. This gene also appears to be up-regulated in bulbectomized mice, which would contribute to cell death in traumatic anosmia. An increase in apoptosis, suggested by increased levels of capsase 3, is also noted in rats exposed to cigarette smoke when compared with normal controls. This increase in the executioner enzyme was noted in the epithelium and nerve bundles in the lamina propria, showing an increase in apoptosis in the cell bodies and axon bundles of mature olfactory sensory neurons. The increase in capsase 3 was also noted in the progenitor basal cells and the supporting cells of the exposed epithelium, which would impact the neurogenesis of replacement sensory neurons.
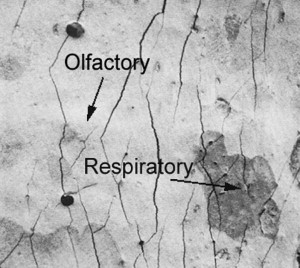
The question remains to what degree the histologic changes seen in the aging olfactory mucosa are due to years of environmental exposure versus other changes in the intrinsic controls of neurogenesis. Rawson and colleagues found that it was just as likely to obtain functional olfactory neurons from the olfactory mucosa of older patients as from younger patients, suggesting that if there is a loss of olfactory neurons in older subjects it is distributed equally throughout the mucosa. Other studies such as that by Loo and colleagues suggest that exposure to the airflow environment may have a role along with changes in neurogenesis seen with aging. In these studies, Loo and colleagues noted a difference in the histologic appearance of the anterior olfactory cells when compared with the middle and posterior cells in aged rats. In older hybrid rats protected from external pollutants, the number of mature olfactory neurons did not change markedly as a function of age in the middle and posterior aspects of the mucosa. The distribution of basal cells in this area was similar to that in younger rats; however, the number of immature olfactory neurons was fewer in the middle and posterior zones of the older animals when compared with the younger rats. The results demonstrate a decrease in the population of immature neurons relative to the population of mature neurons. This finding is in contrast to the anterior-dorsal septal cells, which had a higher density of proliferating basal cells than seen in younger rats. The epithelium in the anterior dorsal aspect was abnormal in several respects, with some areas grossly distorted. The number of immature neurons increased and the number of mature neurons decreased as a function of age. The decrease in proliferating cells in the posterior aspect of the epithelium in aging rats suggests some intrinsic controls on neurogenesis may be unrelated to the rate of neuronal death. The anterior epithelium in older rats showed damaged mucosa with a decrease in mature neurons, disordered mucosa, and an increase in immature neurons. This finding suggests that, in these environmentally protected rats, there may be a role for airflow alone contributing to the changes, because similar findings are noted on the open side of rats with unilateral nares occlusion.
Central Connections
Intact sensation of olfactory stimuli relies not only on intact olfactory receptor neurons but on intact and appropriate synapses as the signal moves centrally. Christensen and colleagues have shown that nerve transection disrupts normal targeting of the olfactory bulb by the immature neuronal cell. Other factors that contribute to peripheral neuron deterioration, such as smoking and viruses, may also ultimately have a role in the development of aberrant synapses and may contribute to the loss of olfactory bulb mitral cells with age. In the olfactory bulb, the number of mitral cells continuously decreases with age. It is estimated that the adult olfactory bulb contains about 60,000 mitral cells at age 25 years and only 14,500 at age 95 years.
Neurodegenerative Disorders
Damage to the cells of the olfactory bulb and central pathways is seen in many neurodegenerative diseases that affect the elderly. Smell sensation deficits occur in many neurodegenerative diseases, such as Alzheimer’s disease, Parkinson’s disease, multiple system atrophy, Huntington’s disease, multiple sclerosis, and amyotrophic lateral sclerosis. The incidence and degree of smell loss in these patients can be variable and often is dependent on disease severity. The characteristics of the smell deficit are dependent on the site of the lesion.
Alzheimer’s disease is the most common neurodegenerative disorder in humans and is the major cause of dementia in the elderly. The olfactory deficits seen in these patients are usually perceptual. Neurofibrillary tangles composed of an abnormally phosphorylated fibrillar microtubule-associated protein (tau) have been implicated as one of the pathologic lesions in the brain tissue of patients with Alzheimer’s disease. The Braak stage has been used to describe the severity of this pathology, with stages 5 and 6 being the most severe. Several studies have demonstrated similar tau pathology in the olfactory bulbs of patients with definitive Alzheimer’s disease. This tau pathology is seen early in the disease process and increases with the severity of the disease. Clinical dementia is highly correlated with both Braak stage and olfactory system tau scores. Attems and colleagues have suggested that in the future biopsies of the olfactory mucosa looking for tau pathology may be useful in determining the risk of future cognitive decline. Early stage Alzheimer’s disease can result in a loss of odor detection and odor discrimination as the tau pathology has more of a peripheral effect that is separate from any cognitive decline. Olfactory loss in these patients can be seen as an early feature of the disease. As the disease progresses and more tau pathology is seen in the central olfactory areas, patients ultimately develop odor identification deficits. Individuals who are heterozygous or homozygous for the apolipoprotein E (APOE 4) genotype have a higher risk of developing Alzheimer’s disease. Tsuboi and colleagues showed a correlation between APOE 4 and tau pathology in the olfactory bulb in a gene dose-dependent manner. Nondemented carriers of APOE 4 have worse olfactory identification scores than APOE 4–negative persons. Olfactory dysfunction and APOE 4 are associated with a greater risk of cognitive decline.
Parkinson’s disease is the second most common neurodegenerative disease in humans. Olfactory dysfunction is a significant feature of Parkinson’s disease, and testing of smell identification can be useful in differentiating Parkinson’s disease from other extrapyramidal disorders such as progressive supranuclear palsy in which odor identification and threshold remain intact. Olfactory abnormalities are detected early in Parkinson’s disease but remain stable as the disease progresses. Lewy bodies are the pathologic lesions seen. They are composed of aggregated fibrillar alpha synuclein and are seen in the olfactory bulb and the lower brain stem. Lewy bodies then develop in the midbrain, limbic system, and neocortex. Olfactory dysfunction is similar in different types of Parkinson’s disease. Problems with odor recognition have been noted in patients. Masoka and colleagues noted that patients who had Parkinson’s disease needed a higher odor concentration to recognize an odor. Semantic processing is also lower than normal in these patients.
Other neurodegenerative disorders such as semantic dementia and frontotemporal dementia which have no olfactory nerve or pyriform cortex pathology can show normal odor perception and discrimination but impaired odor identification. This characteristic can point to the central pathology of the temporal lobes seen in semantic dementia, which have a role in olfactory memory, and that of the frontal lobes seen in frontotemporal dementia, which are involved in odor processing. The orbitofrontal areas are implicated in the determination of pleasantness of odors and taste and in the feeling of satiety, which can have a significant role in altering eating behaviors.
Olfactory Flavor
The appreciation of flavor requires an integration of somatosensory and chemesthetic sensation from the oral cavity and oropharynx and intact retronasal olfaction. Chewing warms and releases odors, and the pressure changes created by swallowing pumps the odors through the oral pharynx and nasopharynx to the olfactory epithelium. Duffy and colleagues have shown that, when compared with young subjects, elderly individuals required 49 times the concentration of olfactory flavoring for consistent detection. This loss of olfactory flavor is consistent with the decrease in olfactory function previously described; however, conditions in the mouth may affect retronasal olfaction and the appreciation of flavor independent of olfactory thresholds. Duffy and colleagues showed that dentures that cover the palate impede olfactory flavor sensitivity even with normal olfactory (smell) sensitivity. It is thought that this effect may be due to interference with normal chewing and mouth movements and may affect the mechanics of odor propagation up to the olfactory cleft. Oral cavity health is another variable when one considers the many etiologies of smell loss and the impact on the health and nutrition of the elderly.
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


