Bronchopulmonary C fibers and acid-sensitive, capsaicin-insensitive mechanoreceptors innervating the larynx, trachea, and large bronchi regulate the cough reflex. These vagal afferent nerves may interact centrally with sensory input arising from afferent nerves innervating the intrapulmonary airways or even extrapulmonary afferents such as those innervating the nasal mucosa and esophagus to produce chronic cough or enhanced cough responsiveness. The mechanisms of cough initiation in health and in disease are briefly described.
The cough reflex protects the airways and lungs from aspiration, inhaled irritants, particulates, and pathogens and clears the air spaces of accumulated secretions. Studies in animals provide conclusive evidence that cough is initiated by activation of vagal afferent nerves. Precisely which afferent nerve subtypes regulate cough has been debated and reviewed elsewhere. In this review, the author describes the stimuli that initiate cough and the mechanisms by which these stimuli activate airway sensory nerves. These data are related to the known physiologic properties of bronchopulmonary afferent nerve subtypes. The review concludes with a discussion of the potential interactions between afferent nerve subtypes and the possible mechanisms of altered cough reflexes in disease.
Chemical and mechanical stimuli that initiate coughing
Multiple chemical and mechanical stimuli initiate coughing in humans and in animals. Several of these stimuli, including capsaicin, citric acid, hypertonic saline, and low chloride buffers/solutions are often used to evoke cough experimentally. Other stimuli known to initiate coughing in animals and humans include particulate/dust; mechanical/vibratory stimulation of the airway mucosa larynx or chest wall; chemical irritants, such as resiniferatoxin, cinnamaldehyde, and allyl isothiocyanate (AITC); and the autacoids bradykinin, anandamide, and prostaglandin E2 (PGE2). Although seemingly varied in origin and chemical composition, many of these stimuli share common modes of action. For example, acids/protons, capsaicin, resiniferatoxin, and anandamide all act in part or entirely by activation of the ion channel/receptor transient receptor potential vanilloid 1(TRPV1). Bradykinin and PGE2 may also act in part through TRPV1 activation. Cinnamaldehyde, AITC, and several other known respiratory irritants (eg, cigarette smoke, toluene diisocyanate) activate the ion channel transient receptor potential ankyrin (TRPA1). Knowing the specific ion channels and receptors for the stimuli that initiate cough is important, as this information can be used to identify which afferent nerves express these ion channels and receptors. This information can then be used to identify possible mechanisms for coughing in disease, but may also suggest therapeutic approaches to restore normal cough function and sensitivity ( Table 1 ).
| Stimulus | Mode of Action | Afferent Nerves Targeted |
|---|---|---|
| Capsaicin | TRPV1 | C fibers |
| Bradykinin | Bradykinin B2 receptors | C fibers |
| Acid | TRPV1, acid-sensing ion channels | C fibers, cough receptors |
| Particulates | Unknown | Cough receptors, C fibers |
| TRPA1 agonists | TRPA1 | C fibers |
| Prostaglandin E2 | Prostaglandin EP3 receptors | C fibers |
| Nicotine | Nicotinic receptors | C fibers |
The stimuli that do not initiate coughing are equally helpful when attempting to identify the specific vagal afferent nerve subtypes that regulate this reflex ( Table 2 ). These stimuli may also suggest a great complexity underlying the cough associated with disease. Consider, for example, three of the most common causes of chronic cough: asthma, gastroesophageal reflux disease (GERD), and upper airway inflammatory diseases (eg, allergic rhinitis, sinusitis).
| Stimulus | Reflexes Evoked | Afferent Nerves Targeted |
|---|---|---|
| Bronchoconstriction a | Mucus secretion, tachypnea | RARs |
| Esophageal acid | Bronchospasm, mucus secretion | Esophageal nociceptors |
| Upper airway stimulation | Sneeze, mucus secretion | Trigeminal afferent nerves |
| Inc. airway luminal pressure | Respiratory slowing | SARs |
| Dec. airway luminal pressure | Tachypnea | RARs |
| Adenosine | Tachypnea, dyspnea | RARs, C fibers |
| Pulmonary embolism | Tachypnea, dyspnea | RARs, C fibers |
a Bronchoconstrictors including histamine, methacholine, leukotriene D4, thromboxane A2, and neurokinin. Failure to reliably evoke coughing in humans or in animals despite initiating reflex bronchospasm and mucus secretion, an increase in respiratory rate and dyspnea.
These conditions are often found simultaneously in patients who have chronic cough. Although none of these disorders are adequately described by a single presenting symptom, it is generally agreed that reversible airways obstruction, acidic refluxate in the esophagus, and inflammation of the upper airways are characteristics of asthma, GERD, and upper airway diseases, respectively. Remarkably, however, bronchospasm, acid in the esophagus and upper airway challenges with a variety of inflammatory mediators are consistently ineffective at initiating cough in animals or human subjects. The inability of these stimuli to initiate cough is not because they fail to activate sensory nerves. Histamine, capsaicin, allergen, and bradykinin all initiate sneezing and reflex-dependent mucus secretion when delivered selectively to the upper airways; whereas the bronchoconstrictors histamine, substance P, and even methacholine evoke reflex-dependent changes in autonomic tone in the airways; changes in breathing pattern; and respiratory sensations, such as chest tightness and dyspnea. Esophageal acidification is known to evoke reflex bronchospasm in animals and in human subjects but has rarely if ever been reported to initiate coughing. Observations such as these argue against the afferent nerves responding to these stimuli as the primary initiators of cough in disease, which implies the involvement of other afferent nerves and afferent-nerve subtype interactions in disease. Defining precisely how these afferent nerves interact to produce the coughing observed in diseases, such as asthma and GERD, are likely be critical to the development of better therapeutic strategies for the treatment of chronic cough.
Afferent nerves regulating the cough reflex
Coughing can be partitioned into at least four phases. The initial phase comprises the encoding of action potentials by the afferent nerves directly responding to the tussive stimulus and the subsequent reconfiguration of the respiratory motor drive within the brainstem. This initial phase immediately precedes any change in respiratory muscle activity. The second phase is the enhanced inspiratory effort that accompanies cough. The expiratory phase of cough has two components: (1) an initial compressive phase when expiration is initiated against a restricted or occluded upper airway; and (2) the expulsive phase, when the upper airways are dilated, allowing forceful expiration and the high airflow velocities that facilitate airway clearance.
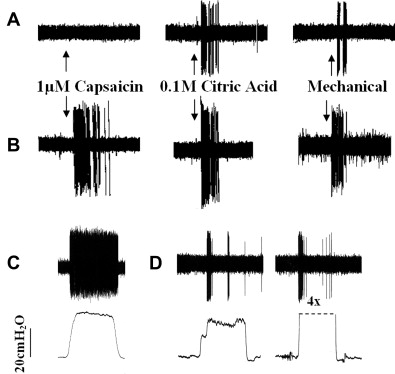
Depending on the stimulus and social situation, the number and forcefulness of the resulting coughs can vary substantially. Given this complexity and the multiple elements involved (lower and upper airways, respiratory muscles, brain stem), it is apparent that multiple afferent nerve subtypes act in concert to regulate the sensitivity, forcefulness, and repetitions of coughs in response to all tussive stimuli. The following discussion focuses only on those afferents involved in the initial encoding phase of cough ( Fig. 1 ).
C Fibers
Bronchopulmonary C fibers are identified physiologically by their action potential conduction velocity, which falls in the C range (≤1 m/s) of the compound action potential. C fibers comprise the majority of afferent nerves innervating the airways and lungs, terminating in the mucosa and submucosa of the nose, pharynx, larynx, trachea, bronchi, and throughout the lungs. C fibers project to the airways from the jugular and nodose ganglia of the vagus nerves, and from thoracic dorsal root ganglia.
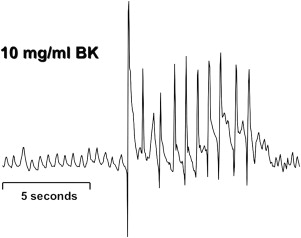
Bronchopulmonary C fibers are activated by capsaicin, bradykinin ( Fig. 2 ), protons, nicotine, and the TRPA1 agonists cinnamaldehyde and AITC. C fibers are selectively desensitized by high-dose capsaicin treatment that also prevents coughing evoked by citric acid. In guinea pigs, C fibers use the peptide neurotransmitters substance P and neurokinin A that act primarily by way of neurokinin 1 (NK 1 ), NK 2, and NK 3 receptors. Neurokinin-receptor antagonists prevent coughing evoked in guinea pigs by capsaicin and citric acid. These results and observations provide conclusive evidence that C-fiber activation initiates coughing.
Despite the overwhelming evidence that C-fiber activation can initiate coughing, the role of C fibers in cough remains controversial. Much of the controversy arises from the inability of C-fiber selective stimuli to initiate coughing in anesthetized animals. It seems likely that anesthesia selectively inhibits C-fiber–dependent coughing. But anesthesia does not readily explain the observation that C-fiber activation not only fails to initiate coughing in anesthetized animals but also actively inhibits coughing evoked by activation of other afferent nerves subtypes. The author speculated that these opposing effects of C fibers on cough might be attributable to the opposing actions of C-fiber subtypes. Studies performed in guinea pigs using stimuli that are selective for vagal C-fiber subtypes suggest that C fibers arising from the nodose ganglia can acutely inhibit coughing, whereas activation of C fibers arising from the jugular ganglia sensitize or initiate coughing. Circumstantial evidence suggests that C-fiber subtypes in humans play similar opposing roles.
Afferent nerves regulating the cough reflex
Coughing can be partitioned into at least four phases. The initial phase comprises the encoding of action potentials by the afferent nerves directly responding to the tussive stimulus and the subsequent reconfiguration of the respiratory motor drive within the brainstem. This initial phase immediately precedes any change in respiratory muscle activity. The second phase is the enhanced inspiratory effort that accompanies cough. The expiratory phase of cough has two components: (1) an initial compressive phase when expiration is initiated against a restricted or occluded upper airway; and (2) the expulsive phase, when the upper airways are dilated, allowing forceful expiration and the high airflow velocities that facilitate airway clearance.
Depending on the stimulus and social situation, the number and forcefulness of the resulting coughs can vary substantially. Given this complexity and the multiple elements involved (lower and upper airways, respiratory muscles, brain stem), it is apparent that multiple afferent nerve subtypes act in concert to regulate the sensitivity, forcefulness, and repetitions of coughs in response to all tussive stimuli. The following discussion focuses only on those afferents involved in the initial encoding phase of cough ( Fig. 1 ).
C Fibers
Bronchopulmonary C fibers are identified physiologically by their action potential conduction velocity, which falls in the C range (≤1 m/s) of the compound action potential. C fibers comprise the majority of afferent nerves innervating the airways and lungs, terminating in the mucosa and submucosa of the nose, pharynx, larynx, trachea, bronchi, and throughout the lungs. C fibers project to the airways from the jugular and nodose ganglia of the vagus nerves, and from thoracic dorsal root ganglia.
Bronchopulmonary C fibers are activated by capsaicin, bradykinin ( Fig. 2 ), protons, nicotine, and the TRPA1 agonists cinnamaldehyde and AITC. C fibers are selectively desensitized by high-dose capsaicin treatment that also prevents coughing evoked by citric acid. In guinea pigs, C fibers use the peptide neurotransmitters substance P and neurokinin A that act primarily by way of neurokinin 1 (NK 1 ), NK 2, and NK 3 receptors. Neurokinin-receptor antagonists prevent coughing evoked in guinea pigs by capsaicin and citric acid. These results and observations provide conclusive evidence that C-fiber activation initiates coughing.



